Instinct: A study in social psychology
Chapter 11:Heredity and the Instincts
Luther Lee Bernard
Table of Contents | Next | Previous
In the foregoing discussion a certain definite relationship between instinct and inheritance has been assumed. This assumption is justified because the accepted definition of instinct describes it as a definite inherited stimulus-response process. Yet to many, perhaps, it may seem that there is an appreciable distinction between an instinct and an inherited trait or unit character. If there is such a distinction it should be possible to define it. To those who accept the Mendelian hypothesis of heredity it is an approved principle that the method of inheritance is by means of unit characters. That is to say, we inherit specific structures, and not merely general qualities. Whether it be eye color, texture of skin, shade of hair, stature, or color blindness, it is a definite thing which we inherit.
What is said here of the inheritance of organs and structures, or unit characters, is also true with regard to the inheritance of instincts. Here the inheritance is of definite, definable groupings of traits, that is, of neural organization. Moreover, this inheritance is always structural, that is, of some physical or biological organization, not of abstract relations or values. To be sure we are accustomed to describe the instincts in a somewhat different way from that in which we describe the unit characters of Mendelian inheritance. In the latter case we are professedly speaking of structure, while ill the former we commonly have in mind the activities which arise as a result of the structural inheritance, neural or non-neural. Thus we speak of the inheritance of the swallowing instinct or of the instincts of crawling and walking.
(249) Here we are referring to an activity. Clearly enough, however, . there can be no such thing as the inheritance of an activity g as a Mendelian or unit character, for an activity is an abstraction, that is, an abstract synthesis of concrete movements, when viewed from the structural side. The term inheritance or instinct used in this way is really a misnomer. What we actually mean to indicate is that we inherit a structure or an organization or a chain of simple structures which enables us to act in a characteristic way with a minimum of trial and error. The instinctive structure should go into action without calling forth a consciousness of effort and without involving a learned readjustment to the stimulus which sets off the characteristic action. This point is well illustrated by the following passage, referred to in an earlier chapter, describing the working of the flying instinct in the fly: "The nervous mechanism is perfected, and when hatched and dry the fly makes its first circuit as accurately as if it had practiced the movement for days."[1] This initial perfection of the instinct is denied, as was brought out in the discussion in a previous chapter. But, as was there indicated, the denial is justified only when the instinct itself is incompletely organized, as is so frequently the case in the higher or habit-forming animals. In the lower forms of animal life the initial accuracy of the instinct is much as Gamble has here described it. But in the higher forms the instinct becomes distorted, either through being bred out by the substitution of environmental controls for internal organization and control, or because of the lengthening of the period of infancy, which displaces the instinct organization and prevents it from functioning in unmodified form. That is, habits are already formed before the instinct gets into operation. In the former case, we have vestigial instincts; in the latter, we have delayed instincts. In the one case the disrupted or atrophied instincts may be
(250) reconstructed as habits by organization and training; in the other the instincts are likely to be repressed or considerably modified by the habits which are formed before the instincts have a chance to come to maturity.[2]
It is only through conceiving of instincts in terms of definite structures that we can reach a definition or description of them which is more than merely a logical arrangement of word symbols which may or may not have any accurate relation to structural facts. The futility of speaking of activities or attitudes as instincts, when these activities and attitudes cannot be reduced to unit structural organizations capable of transmission by means of biological heredity, at once becomes apparent. It is not enough, however, that we find the gross physical structure or the inner neural correlate of an activity or attitude in order to establish the fact that it is an instinct. Such a structural organization may be an acquired habit, for habits must have their physical and neural organization bases quite as well as instincts. It is true, nevertheless, that there is no instinct where this unity of structure does not exist. We must learn to distinguish between the inherited and the acquired nature of such underlying physical organizations. The final criterion here is one of fact. That is to say, was this organization determined in the chromosome structure of the reproductive cells of the parents of the individual, and did the individuals themselves, as a matter of fact, develop this organization without making a trial and error adjustment and without environmental determination exercising a predominating influence? Or, did the activity in reality come from a modification of structure environmentally induced, either consciously or unconsciously; postnatally, prenatally, or preconceptually ? [3]
( 251)
These questions can be answered finally only through the aid of a mass of detailed investigation which now has only been begun. They certainly cannot be settled on the mere basis of opinion or through speculative or a priori reasoning. We can, however, aid in clearing away some of the obstructing debris of prejudice, dogmatism and opinion in regard to these matters by invoking a definite statement of the facts of heredity as they bear upon the disputed question of the relation of instinctive to acquired activity. If we can show the biological limits and possibilities of inheritance, on the one hand, and if we can make clear the conditions under which characteristics can be acquired and what types of characters may be acquired under these conditions, on the other hand, we shall then and only then be in a position to begin the process of gradual accumulation of facts. Only through working out such a method of approach shall we be able to see clearly where our facts lie and to define and classify them when we find them. In the present and subsequent chapters such a delimitation of the field of heredity will be attempted.
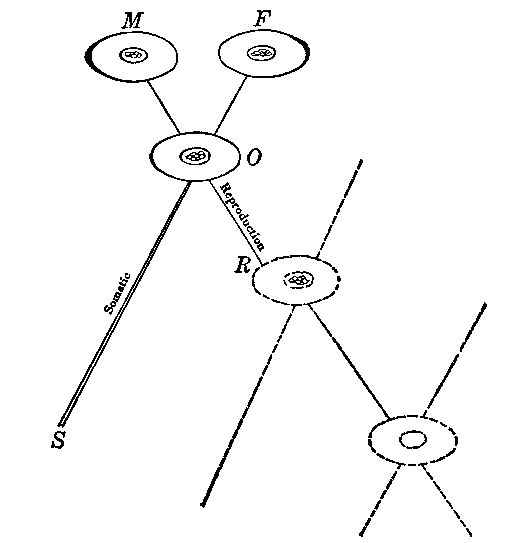
The new individual arises from the union of the parental reproductive cells. This fact may be diagramed symbolically as follows, it being kept in mind that the diagram is only a symbolic, not a photographic, representation. M and F symbolize the male and female reproductive cells respectively; O the offspring cell resulting from fertilization; and the lines OS and OR symbolize the development, through division, of the somatic and reproductive cell aggregations respectively. Detailed descriptions of the mechanics of fertilization and heredity and of the growth of the organism may be found in the standard works on these subjects. The offspring begins its existence as a single cell resulting from the union of the matured parental cells, including cytoplasm as well as chromosome content. The future development of the new individual comes through a process of cell division. This cell division
(252) in the somatic tissues of human beings proceeds to a remarkable extent, the multiplication of body cells aggregating something like 26,500,000,000,000.[4] The number of reproductive cells differs for the male and female. Lode calculated that
approximately 340,000,000,000 spermatozoa are developed in the male between the ages of twenty-five and fifty-five.[5] But the new born female contains only 36,000 to 200,000 ova, "of which no more than about 30,000 remain at puberty, the others having undergone atrophy and having served as food material fur the more vigorous ova. Of these 30,000, no more than about 400 become mature, one at each menstrual period between the ages of about fifteen to forty-five."[6]
(253)
Although the new organism begins its separate or individual existence as a single cell, this initial cell divides into two general types of cells, the somatic and the reproductive, as was implied above. The reproductive cells are highly specialized and do not perform any adjustment function with regard to the individual's environment. The reproductive cells are split off from the somatic cells in the first few hours of the existence of the new organism, the separation occurring among the first cells which result from the division and multiplication of the new individual cell organism. The result is that the reproductive cells are very early removed from the possibility of being easily contaminated by the environmental accidents of the somatic cells, since they are split off before the somatic organization has had much opportunity to be influenced by environment. Ever after this initial separation the reproductive cells maintain their separate existence and reproduce themselves by fission, just as do the somatic cells. They are segregated and stored in an inner portion of the body where they will be as free from external influence and harm as possible. They are normally discharged for reproductive purposes, though the discharge mechanism is not so definitely and instinctively adjusted to stimuli connected only with reproduction that the cells are discharged either in the most economical or effective manner or even solely for reproductive ends. Nature being blind, as the old aphorism has it, is prodigal here as elsewhere in order to make sure of some, though possibly not the best, results. This fact is abundantly testified to by the vast excess of spermatozoa over ova. The hereditary determiners are organized in the chromosome content of the nuclei of these reproductive cells. The nucleus or inner portion of the cell is separated from the external portion of the cell or cytoplasm by means of a surrounding sac or tissue. Although there is interchange
(254) between the two parts of the cell, the inner portion is more protected from outside influences and leads a more isolated existence. It carries the chromosome content which determines the inherited traits of the offspring. The function of the reproductive cells, apparently, is to carry these hereditary determiners of the chromosomes as safely as possible and as completely unmodified by the somatoplasm and external environment as possible and to aid in their deposit under such conditions as will secure fertilization and initial growth. An elaborate bodily structure makes possible this process.
As stated above, this inheritance is apparently determined through the chromosome content organized within the nuclei of the reproductive cells. Some writers on biological subjects speak of the possibility of cytoplasmic inheritance,[7] but the prevailing opinion appears to be to the contrary. The current conception of heredity is that it is a matter of chromosome determination, while influences from cytoplasm are perhaps more properly classified under environmental factors. Julian S. Huxley is of the opinion that the majority, possibly all, characters are due to the Mendelian genes and finds no evidence to the contrary.[8] He cites approvingly Morgan's view [9] that each chromosome represents a linkage group of characters and that the intensity of the linkage varies inversely with the linear distance of the genes along the chromosomal frame-work. The theory is that the chromosomes which determine the heredity do not act as individual units in this determination, but that parts of the chromosomes operate as individual factors.[10]
( 255) Huxley adds, "The chromosomes and their contained genes may be looked upon as the regulating mechanism which makes heredity, and indeed any real organic constancy from generation to generation, possible at all— the regulating mechanism of the species . . . . The gene-constitution of the organism is a mechanism for preserving the character, number, proportions, and locations of a great number of unit factors; and it accomplishes this by being itself a unit of a higher order, whose parts are linked together in a definite and specific way, each of the parts, and therefore the unit-complex as a whole, being self-perpetuating." [11]
This mechanism for controlling the heredity or making for the continuity of the type is chemical in character. It also resists modification from the external environment, although it is sufficiently susceptible in the long run to the influence of some external agency that the character of the heredity or of the inherited organization may be modified.[12] Some think that we may even learn in the course of time to alter the genes at will and thus direct the heredity in the channels which we desire, but there certainly is no immediate prospect of this.[13] Huxley thinks that the only possibilities of such direct modification are, (1) that some side-chain of a gene may be so altered that the details of its working will be changed while the main essentials remain the same, and (2) that by stimulating the gene to overproduction the structure or content of the inheritance determiners may undergo change.[14] R. S. Lillie is of the opinion that possibly "the appearance of a sudden variation or mutation in an organism is the result of a chemical change," such as the substitution of one chemical for another in the organism on the analogy of substituting cadmium for
( 256) zinc in the growth of physical objects, by which changes of form and structure are induced.[15] He continues, "The formation of a new compound in formative metabolism may thus mean the appearance of a new structural and physiological character."[16]
Environmental factors operate primarily in modifying or conditioning the growth of the organism, rather than in directly determining its heredity. Especially, acquired characters are not transmitted by inheritance, except possibly in relatively rare and poorly understood instances, such as in that of the much discussed case of Guyer and Smith's experiments on rabbits.[17] While the influence of the environment in a non-inheritance manner is shown where outside factors produce artificial fertilization of the eggs,[18] growth in the organism apparently is produced by the genes themselves. Some factor leads to the production of certain specific ferments, and different multiple allelomorphs of a single factor may represent different degrees of oxidation, while other factors may lead to the production of substances which inhibit
( 257) the action of these ferments, and growth is controlled accordingly.[19] "Whether these ferments are produced directly by the individual gene, or as the end-result of a long series of reactions in the cell, we have as yet no means of telling." [20] Thus, the differentiations of the developing fertilized egg take place as the result of the reaction of the genes with the environment. The cytoplasm influences development, not in determining the character of the differentiation, but as a basic material necessary to the life and the complete development of the cell. The organ-forming stuffs in the cytoplasm of the egg aid in the early development of the organism.[21] The differential development of the organism, in early stages, and otherwise, is determined largely by the organization of the environment in the large, which selects the gene-produced structures and organizations which are fitted. Some cells are immune to certain types of environmental selection and others are susceptible. Also, environmental control mechanisms, such as endocrine secretions, play a large part in selecting the direction of the growth of the organism.[22] The material of the gene probably in some cases actually corresponds with the material of the adult organ which it helps to determine.[23]
The somatic cells multiply through division into aggregations which are of four general hereditary types. The total conformation and organization of these somatic cells develop according to general organic type. The somatic cells splitting off from the initial offspring cell, resulting from the union and fertilization of two reproductive cells, from dogs, for example, always aggregate in the form and structure and chemical composition and organization of dogs and not of other animals. The same is true fir all ether animals and for plants. In like
( 258) manner, these differentiated somatic cells aggregate into the form of those organs or unit aggregations of cell structure within the total structure or organism which normally characterize the type. Each animal has a head and a head of a certain type according to that variety, species, genus, etc., to which it belongs. These two types of hereditary aggregations of somatic cells are closely correlated in their development. A third type of aggregation of somatic cells is also noticeable: the development of differential traits, such as eye, hair, and skin color, head-shape, nasal and facial indices, and the like. These are the traits to which we commonly apply the Mendelian principles of inheritance.
A still more specific and detailed structural inheritance is to be found in the nervous system. Just as the general and specific physiological and anatomical organization of the organism is inherited through the chromosomes, so various elements of neural structure or organization, which serve the organism in making rapid and accurate adjustments to its environment,[24] are inherited. These are the reflexes and the instincts. They have come into the inheritance process by natural selection in the service of survival, or otherwise, in the same general way in which the other factors of inheritance arose. It is reasonable to suppose that they are subject to the same laws of origin and transmission. They differ merely in being primarily facts of neural organization, with their end organs and muscular and glandular response connections, rather than of muscular and other largely external and gross tissue organization and chemical content and distribution. They may also differ in complexity. But they, equally with the others, belong in the category of inheritance and are subject equally to the laws of unit characters. All of these traits are inherited, that is, they come down through the
( 259) structures and the chromosome determiners of the reproductive cells and are not acquired from the environment. The chromosome filaments of the two parent cells uniting in the new derivative cell apparently control the organization and distribution of the somatic cells, which arise through division, and group them according to type.
Four facts having definite bearing on the hereditary and unitary nature of the instincts should be noted in connection with the fertilization process and the development of the somatic cells. First, the hereditary equipment of the new-formed individual is completed at the point of fertilization. Any characteristic received after the process of fertilization has taken place, that is, not contained in the chromosomes of the uniting parent cells, is not inherited. It is acquired. Likewise, any predisposing influence which affects the outer portions of the parental reproductive cells before fertilization, but which does not modify the determiners or chromosome content, cannot be classified as an inheritance or instinctive influence or factor. It belongs to a category which we may, for want of a better term, denominate pre-conceptual environment. The nature of the new individual, so far as his inheritance is concerned, is formed once for all at the point of fertilization, where the chromosomes unite and determine their control over the newly forming somatic organization. "The impregnated egg cell in the process of its first division into two new cells typifies the entire development of the completed human being. There within that minute compass lie the materials which predetermine with fatal exactitude the future of the adult form, its sex, its type of stature, the blending of the traits which it owes to its parents and their parents."[25] Many more characteristics will be acquired by the
( 260) somatic constitution from the various environmental pressures, and, with regard to some types of organization, the individual may even be primarily made over as contrasted with the line of development indicated in his inheritance determiners or chromosome content. But this acquisition of a made-over content is not brought about through inheritance. It comes from the environmental factors working upon the inherited equipment and modifying it. Of course there are very obvious and definite limits within which the hereditary equipment can be modified. One cannot transform by any sort of environmental pressure one type of animal into another or one organ into another organ or, possibly, black hair into red hair. It is possible, however, to train one animal or one organ to perform in large measure the functions of another, or artificially to color hair. Also various types of intelligences or mental coördinations can be built up on the basis of inherited neural organizations or dispositions. In the matter of moral and social attitudes the individual can be almost completely transformed through environmental pressures, because such characters are not in the first instance inherited, or at least the inherited basis is not usually rigidly determinative of the moral and social traits which are later acquired.
(a) Only the structural organization of the somatic cell aggregations is determined by the hereditary determiners in the new cells through the fertilization process. Somatic structures of the sort described above are inherited directly (unless acquired from environmental pressures or conditions). Mental, moral and social traits are not thus predetermined in the chromosome organization in the same manner as the simple somatic traits or organic structures are; for the former, owing t0 their abstract quality, d0 not represent any fixed relation to physical structure at all. Therefore, they are not inherited. There is no way in which these highly variant and variable traits could be directly determined by the chromosome con-
( 261) -tent, for the process of inheritance determination is biological, while the determination of these traits is at least partly social. It is true, however, that the somatic structures, including the gross organic structures and any preformed neural dispositions upon which the mental, moral and social characteristics rest, are thus determined in the chromosomes. In turn they largely determine, indirectly, by means of their functioning, the non-physical abstract or ideational characteristics. Thus one might be inclined to speak of an indirect determination or inheritance of mental, moral and social characteristics. Such, however, would be incorrect; for the determination of these abstract traits from the standpoint of heredity is only partial and often minor. Subsequent chapters will undertake, in some slight degree at least, to indicate the extent to which environing factors determine these nonphysical traits. It is because of this necessary distinction between inheritance and environmental factors in the determination of these mental, moral and social traits that we find it inaccurate to speak of them as instincts in the same sense that we speak of the activities based wholly on inherited somatic structures as instinctive.
(3) It should also be further emphasized that the somatic structures do not influence the inheritance determiners of the reproductive cells in any specific way. This is a fact which is generally accepted. The reproductive cells are completely surrounded by the somatic cells and draw their sustenance from them and are therefore subject to general influences of various sorts from them. But these influences are external rather than in the nature of specific modifications of the chromosome contents. Various toxic conditions in the somatic organization may he carried over into the reproductive cells and produce changes there, such as disintegration, abnormally stimulated development, sterility (because of injury to the cell content), or even destruction. But no machinery
( 262) is known by means of which a specific character of the somatic organization, inherited or acquired, could conceivably impress itself upon the inheritance determiners, "photographically" or "telegraphically" as it were, in such a way as to cause the inheritance of that same trait in the offspring. Because of the improbability or even impossibility of such direct internal modification of the inheritance determiners we are accustomed to say that acquired characters are not inherited. It can readily be seen from the preceding discussions that the transmission by inheritance of the moral and social complex mental traits acquired by the parents would be even less conceivable or probable than that of the more concrete acquired physical structures and habits.[26]
But if the inheritance of acquired traits— particularly of the mental, moral and social sorts— should, by some unexpected turn in the evidence and experience, be demonstrated, the claims of the environmental interpretation would be strengthened thereby. This fact would establish the direct determination of the instincts by environment, the character
(263) of the environmental pressures impressing itself more directly upon the resulting heredity. According to the currently accepted theories of heredity and environment, the determination of the instincts and other forms of inheritance is indirect rather than direct. Furthermore, there is no copying of the form or type of the environmental pressure into the inheritance mechanism and the resulting hereditary traits, as there might be if the latter were directly determined by the environment. By means of the indirect method of determination the inheritable traits which appear in the organism through the processes of variation or mutation are selected out of the heredity and lost, or selected in and perpetuated, as a phase of the racial adjustment of the type, to which the individual organism belongs, to the demands of its environment. Nothing can survive in the type which is markedly harmful to the individual, because of the advantage in the competitive struggle to exist and reproduce which those individuals not adversely incumbered have over those who are thus handicapped by unfavorable traits. Thus, in any case, it is environment which in the long run determines the inheritance, whether it produces the variations and mutations— the traits— by direct action or merely by indirect or selective pressures. The weight of evidence, however, is to the effect that this environmental determination is indirect and selective rather than direct and formative. It is on the basis of this evidence and assumption that the arguments in this chapter are set forth.
(4) This argument for the non-inheritance of acquired somatic traits is further re-enforced by the fact that reproductive cells do not transmit the same traits as those contained in the aggregate somatic structure. This is equivalent to saying that the hereditary determination in the reproductive cells of an organism is not the same as the visible characters produced through the hereditary aggregation of
(264) the somatic cells of that organism. According to the Mendelian ratios the two series of traits may be represented by the following equations. The results of the original experiment of Mendel are here presented. The letters T and D represent respectively the tall and dwarf strains of peas which
(1) TT x DD= TD TD TD TD (Inheritance)
TT TT TT TT (Somatic traits)
(2) TT x TD= TT TT TD TD (Inheritance traits)
TT TT TT TT (Somatic traits)
(3) TD X DD = TD TD DD DD (Inheritance traits)
TT TT DD DD (Somatic traits)
(4) TD X TD = TT TD TD DD (Inheritance traits)
TT TT TT DD (Somatic traits)
he bred to each other. In the above equations T is dominant over D. In the terms to the right of the equations the symbols above the line represent the distribution of hereditary traits in the unmatured reproductive cells of the four hypothetical individuals proceeding from the union indicated to the left, while the symbols below the division lines represent the traits as they appear in the aggregated somatic structures of each of the four individuals, that is, as they look to the eye. It is interesting to note, in this connection, that social traits are somatic.
The bearing of this fact of the non-inheritance of gross or aggregated somatic characters upon the problem of instinct is that it removes the only substitute possibility of the inheritance of complex mental and moral and social characteristics. If it be proven that these general non-physical train are not determined in the regular inheritance process, it might still be claimed by those who hold to the doctrine of the inheritance of acquired characteristics, as indeed it has often been maintained in the past, that these traits once
(265) acquired may be transmitted biologically. This residual explanation, however, becomes untenable, once it is shown that the gross or visible somatic characters, resulting from external aggregation, are not necessarily the same as those carried in the determiners of the chromosomes. Unless acquired somatic characters can impress themselves directly upon the chromosome content and thus become determinative in the reproductive cells— which, as pointed out under (3) above, is shown by experience not to be the case— they cannot therefore enter into the heredity. It was once supposed that, externally considered, like produces like, that is, that an external characteristic, such as color or stature, in some mysterious way necessarily reappears in the offspring. The distinction illustrated by the above diagrams between the somatic or external traits (those below the dividing line) and the traits implicit in the chromosomes of the cells (those above the dividing line) was not then realized. But there is now abundant evidence to show that inheritance is only through the determiners of the reproductive cells (the traits above the line). Many, including some of those who understand the Mendelian theory as well as those who do not, still fall into the error of speaking of inheritance as if such a reproduction of external traits occurs. Such not being the case, there is at least no more reason for supposing that an acquired somatic trait will reproduce itself through inheritance in the offspring than that an inherited somatic trait will necessarily so reproduce itself.[27] The inherited somatic trait is itself determined by the contents of the cell. That is, it is the end of the inheritance process, and not a link in that process having power to determine links further on. Much less, then, should we suppose that the acquired somatic trait,
(266) which is not even an end link in the hereditary chain, should have the power of influencing the chain of heredity.
In an earlier part of the present chapter, in discussing the criterion for distinguishing between inherited and acquired traits, it was stated that the test resides in a factual determination of whether the activity or attitude sprang up spontaneously and as a result of the natural growth processes, without marked trial and error adjustments, or whether it obviously had to be learned. It is now desirable to state this distinction more specifically and completely. Some activity and attitudinal traits may appear seemingly spontaneously, without obvious learning, and yet they may not be determined in the inheritance process. We may outline the sources of activity as follows:
I. Structure and organization inherited (fixed by the inheritance determiners).II. Structure and organization not inherited.
1. Determined by purely organic conditions of development.
a. Where a developing organ or organization, itself, possibly, but not necessarily, determined by heredity, in turn determines the direction or degree of development of a structure or organ not itself determined in its final form by heredity.
b. Where a structure, whether inherited or acquired, is modified and in some degree determined by mechanical and externally arising environmental conditions.
2. Determined by a more or less consciously and visibly controlled adaptation process.
It is quite clear from the above analysis that all activities arising from adjustment structures and circumstances under
(267) II, 1, a, would appear to superficial observation as inherited. Likewise those activities arising from II, 1, b, where the "accident" occurred before birth, and frequently even after birth, if the connection between the resulting structure and activity had been overlooked, would give the same impression. Such activities, especially those of the former class, would arise automatically with little or no trial and error adjustment. Therefore they could not be distinguished by uncritical observation from inherited acts and attitudes. Consequently we see again illustrated the necessity for a critical analysis, going beneath the surface aspects of phenomena, in order that we may make a dependable classification of instinctive and non-instinctive activities. Unfortunately the experimental biologists are not yet able to give us extensive or completely satisfactory data as to what structures are the result of direct hereditary determination and what ones proceed secondarily from the conditioning pressures of the development of other structures and those arising from external preconceptual and prenatal conditions. However, this limitation to our knowledge does not rear an insuperable barrier to drawing significant conclusions with regard to the respective fields of instinct and acquired habit.