The Fundamental Laws of Human Behavior
THIRTEENTH LECTURE
Max Meyer
Table of Contents | Next | Previous
A simple sensory excitation bringing about a temporally complex response. Some, but by no means all, temporally complex responses to a simple sensory excitation explainable on the basis of geometric-mechanical rivalry of motor organs. Multiform variation of response. Importance of kinesthetic sensory activity. Two stages in the development o f speech. Reflex pointing and utterance of a dental sound. Left-handedness during the first months of life. Right-handed reflex pointing. Right-handedness and speech. Accented and gesticulating languages.
WE have discussed the fact of a temporally complex stimulation, a word, producing one response at a definite motor point. But the reverse is of no less importance, —the fact of an absolutely simple sensory excitation bringing about a temporally complex response, for example, since we are just discussing speech, bringing about the pronunciation of a word, or of a whole sentence. We must not imagine, however, that the pronunciation of every—even the simplest monosyllabic word presents a problem of this sort. When a child one year old begins to pronounce such syllables as ga or da, nothing is required to bring about the proper motor activity but an equal, or about equal, division of the simple nervous process into several branches simultaneously. This is theoretically nothing new, a pure case of
(169) sensory condensation. The muscles active in such a case in producing the second sound, the vowel, may be innervated at the same time with those active in producing the first, the consonant. Everyone can easily make the experiment which proves this. Get ready with your mouth organs to say ga, but stop short before the expiratory "explosion." Then do the same for the syllable goo. You notice a great difference of position of the mouth organs and of tension of the various muscles, although the consonant beginning the word is identical in both cases. The mouth organs, it appears, are ready at once to produce both sounds, consonant and vowel; and it may be altogether a matter of geometric-mechanical conditions an organ being incapable of moving at once in two opposite directions—that actually the consonant, since it cannot occur simultaneously, precedes or follows the vowel. Try the case of the vowel preceding the consonant. Get ready to say og or ot. Again the muscles, especially those of the tongue, have entirely different tensions before even the vowel o is heard; that is, the muscular tensions needed for the second sound begin at the same time as those needed for the first. It is possible—indeed probable—that in the pronunciation of such words as god and dog there is no difference at all in the temporal order of the nervous activities involved, but a mere distribution of the relative resistances of the nervous branches serving simultaneously as conductors, to the effect that, in the one case, the muscular "g-tension" is stronger than, and thus becomes outwardly effective before, the "d-tension, " in the other case the reverse,—the o-tension being of intermediate intensity in either case.
Let us admit that this is the true explanation of the pronunciation of many a word, but let us also be aware that this does not include the assertion that, whenever
( 170) any word is pronounced, its pronunciation results just in this way. Nothing can be more misleading than the tendency to believe that a certain event, in the complexities of human life, must regularly have the same cause. There is no question that the pronunciation of a long, polysyllabic word—not to mention a sentence can not be the outcome of a purely geometric-mechanical rivalry of the several speech organs crowded together in and about the mouth and excited with different intensities. If a short word often does come about by this rivalry, it often may and does come about in the way typical for long words and sentences. What is this latter way?
Let us imagine, in the following discussion, a child so far advanced in experience that he responds to any simple speech sound impressing his ear by producing a similar sound with his own speech organs. In Figure 44, Sk denotes (k equals kinesthetic) a sensory point within the
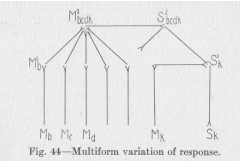
muscular and tendinous organs of speech, or, rather, that whole group of sensory points which are stimulated by the tensions of the speech organs during the production of any definite sound, say, the word baby. The motor ending of the reflex arch Sk Mk is of no direct interest to us; we might imagine that Mk represents the motor activity of saying "baby," which leads again to stimulation of
( 171) Sk, but we must note that in the following figure another point, Ma. will be supposed to take this function. We know that from Sk over a higher center, S2bcdk M2bcdk, a route of low resistance may be established in the direction of numerous motor points, Mb, Mc Md, etc., each of which represents here rather a group of motor points active in the production of one definite speech sound than a single point. To take a concrete example, a child happens to say "baby" and the mother's answering "cry," "hungry," "tired," etc., causes the child to pronounce one of these same words in succession to "baby." While the word baby is—probably repeatedly produced, the kinesthetic nerve process starts from Sk. At the same time the auditory process (not represented in the figure) resulting from the mother's answer leads to the imitative motor activity at either Mb, or Mc, etc. Of these two processes, the former is attracted and deflected by the latter, and the child, instead of saying once more "baby," says now "cry," another time "hungry," etc., whatever the mother has said. The deflection of the nervous process originating at Sk, from S1k upwards (in Figure 44), leads then to a multiform variation of response. The actual response it any future time, on the basis of this variation of the nervous path, to a stimulation of Sk occurs either at Mb, or at Mc, or at Md, etc., according as adventitious circumstances happen to favor the one or the other of these points. That the variation must be multiform, is plain enough, for many kinds of word successions having the same first word must needs occur in the experience of a child surrounded by older people. What, then, may be such an adventitious circumstance as just mentioned, making the selection among the motor outlets of the multiform nervous path? Here we approach the real problem set before us at the end of the previous paragraph.
(172)
Figure 45 is an elaboration of the directly preceding figure. S� is a sensory point, at which, we suppose, an absolutely simple nervous process originates, for example, the sound of a baby crying at a distance. At Sl a branching of the nervous conductor brings about a division of the process to the left and right. We suppose that the resistance to the left is less, so that the main flux occurs in this direction, much less to the right, as indicated in the figure by the conductors to the left being drawn in
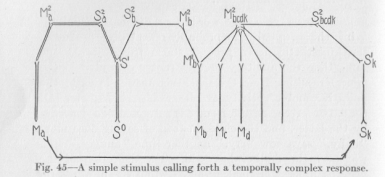
double lines. The motor response occurs practically only at Ma, notwithstanding a certain amount of muscular tension elsewhere, especially at Mb. Now, imagine both these points, Ma and Mb, to represent two groups, partly overlapping, of muscular organs of speech. The sound actually pronounced is then the one corresponding to Ma; the other, corresponding to Mb, is excluded by geometric-mechanical rivalry, the same organ being incapable of moving in two opposite directions and following, by necessity, the stronger pull,—a very ordinary event in animal life. The pronunciation, through its muscular tensions, stimulates a whole group of the kinesthetic sensory points in the organs of speech; and
(173) the group of sensory points thus affected is exactly, let us say, the one which we have formerly called Sk. Then a second word must be pronounced in response to the stimulation of Sk; and of the many responses possible, at Mb, or at Mc or at Md, etc., the one must actually occur which is favored by the adventitious circumstance that directly before this moment a nervous flux, although weak, has passed from S1 over S2b and M2b through the neuron M1b Mb. That is, under the conditions embodied in Figure 45, a simple stimulation of S� must result in a temporally complex response, a succession of two muscular activities, first at Ma and then at Mb.
Still the question is left if in our demonstration just completed we had a right to pre-suppose such a branching of the nervous conductors as represented by Figure 45 at the point S1, which means that saying "cry" (Mb) is a rival, but weaker, response to a certain stimulation at S�(e. g., a sound coming from a distance) that is actually responded to by saying "baby" (Ma). We had a right to pre-suppose this for the following reason. The child in question is sufficiently advanced in experience to respond to a simple speech sound heard by pronouncing it imitatively,—this we have assumed. We only have to ask, therefore, whether he has the opportunity of hearing grown people say both "baby" and "cry" frequently while a crying baby impresses the sensory point S�. Of course, this must be the case, and surely, too, the word "baby" is under these conditions more frequently used than the word "cry," so that of the two paths establishing themselves from S1 upward the one leading to Ma must have its resistance reduced much more than the one leading to Mb.
We have shown, thus, how experience can train the nervous system in such a way that one simple stimulation
( 174) is responded to by a succession of motor activities, by a temporally complex action. We do not assert that every training of this kind muss; conform with figure 45. There may be hundreds or thousands of other possibilities of bringing about a similar result. To have demonstrated one such possibility, as an example, is sufficient for our present purpose. Let us use this opportunity, however, to point out a fact of much general importance for all training of temporally complex reactions, namely, the necessity of kinesthetic sensory activity. If it were not for temporally complex reactions, kinesthetic sensory points would be rather superfluous. Why should any animal respond by particular reactions to tension in its muscles and tendons? The biological purposes of activity are, mainly, protection and nutrition. Neither is, as a rule, served directly by an animal's reacting upon internal tension. But a definite temporal order in a very complex reaction could hardly be acquired, as Figure 45 shows, without the mediation of kinesthetic sensory points, enabling the first motor activity to determine—in co-operation with further conditions the second, the second to determine the third, and so on. The greater the complexity of an animal's anatomy, the greater is the need of temporal order in its compound motor activities. Thus, indirectly, the kinesthetic sensory points come to serve the purposes of nutrition and preservation, to be ultimately as indispensable as the sensory points on the surface of the body. But a direct reference to any purpose can hardly ever be found in the kinesthetic sensory activity, except where—as it happens especially in the highest animal, in man—the performance of a skillful motion becomes itself a purpose, severed from all "practical" significance, for example, in athletic sports.
On the basis of our views concerning the nervous conditions of speech functions, we may adopt a distinction
( 175) which is often made by those interested in the growth of speech. We may distinguish two stages in the development of speech, the early prattle of a child, consisting mainly of monosyllabic words, and the more highly developed—often, but only with partial justice, called exclusively the "imitative" formation of longer words and sentences. The early prattle of a child depends on a branching of the nervous process so that numerous motor points receive the excitation simultaneously or almost simultaneously. The branching of the nervous conductor is then similar to that in the special process of learning which we have called sensory condensation. But this branching may be completely inherited. The speech organs usually produce both a consonant and a vowel, but one precedes the other owing to geometric-mechanical rivalry. Even sounds consisting of two consonants and a vowel may be produced in this way, like gook. And polysyllabic words too: very easily reduplications like dadadada, or words like goocka, booppa, hoppa. Such words are characteristic of the second year of life. It is a mistake to think, however, that the term imitative is in no way applicable to this stage. In this—in the main monosyllabicprattle, as in his later 'talking, the child is influenced by what lie hears grown people pronounce.
The second stage, from the end of the second year continuing really all through life, is governed by kinesthetic sensory fnnctions, as illustrated in Figure 45; and if, as often in the nervous disease of aphasia, the kinesthetic functions are interfered with, the grown person retires to the prattling stage. Let no one think, however, that the simpler function, governed by geometric-mechanical rivalry of the speech organs, has ceased to exist in the second stage. Many kinds of lapses of speech in grown people and in children prove the contrary. When a child
( 176) says tome instead of come or dood instead of good, it is not necessarily, as one might be inclined to believe, because he can not. pronounce gutturals, they are the class of consonants most easily produced. Most probably, in such a case, the nervous function described in Figure 45 is not perfected, and simultaneous innervation, by means of a mere branching of the nervous conductor, governs the pronunciation. Then we understand why tome should be pronounced instead of come, for the branching of the nervous conductor is more ready to serve t and m together, both of which are produced by the speech organs located in the front of the mouth and co-operating naturally in many ways, than to serve k and m together, of which the former is produced by speech organs located in the back part of the mouth and therefore less naturally co-operating with the other. That any imaginary difficulty of producing guttural sounds need not have anything to do with the case, is demonstrated by the example of a German-speaking child who insisted upon saying kragen instead of tragen, certainly not because gutturals are more easily pronounced than dentals, but because the other significant consonants of the word, the German r and the g, are both gutturals, with a common nervous conducting apparatus.
In our ninth lecture we mentioned the reflex of pointing with the index finger at a thing which impresses the eye. We said that this reflex appears at about the same time when the first articulated sounds (usually guttural and dental ga and da) are instinctively produced by the baby. But the act of pointing is not accompanied by a guttural, but by a dental sound the baby pointing and saying dadada. We recall, too, the interesting fact that in all Germanic languages the demonstrative pronouns begin with a dental sound. Why does the baby say da
( 177) when pointing at anything, and not ga? The answer is to be found in the same considerations which we have just applied. The act of pointing sets in motion one of the extremities of the body. The nervous process, one of whose divisions goes into this extremity, can naturally reach by one of its other divisions more easily the muscles of the frontal part of the mouth, belonging to the periphery of the body, than the muscles of the throat, belonging to a different, an internal, system of muscular activity. Try yourself to accompany a pointing movement by a dental or a guttural sound. The latter seems less natural.
When the reflex of pointing first occurs, we notice that the pointing is done far more frequently with the right than with the left hand, whereas previous to this time the right hand is by no means favored in action. The functional connections between the reflex of pointing, the growth of speech, and the development of right-handedness, are so interesting that we must discuss them more in detail. A baby two or three months old, in using the hand, for example in order to put the fingers into the mouth, unquestionably gives preference to the left hand in about two-thirds of all cases. That is, one observes during the first few months about two activities of the left to one of the right hand. Toward the middle of the first year this preference disappears, and both hands are now used with about equal frequency. This is also the time when speech sounds, although hardly yet imitative, become more frequent. During the second year say, at the fifteenth month, making allowance for the enormous individual differences—the right hand begins to predominate, and about the same time speech enters upon that rapid development which insures to this art its being the distinguishing feature of a human being as compared with an animal. As little as it possesses
(178) speech in the human sense, can any animal be said to be right-handed or right-sided. In animals both sides of the body function about equally.
It is probable, although not absolutely certain, that the totality of these facts may be explained in the following way. The left hemisphere of the normal human brain, as has been known for nearly a century, has functions different from those of the right hemisphere; not merely in so far as the right hemisphere is more closely connected with the left side of the body and the left more closely with the right side of the body, but through its significance for the functioning of the speech organs. Certain nervous paths, most probably those leading from the kinesthetic sensory points of the speech organs, have their higher centers in the temporal part of the left hemisphere exclusively, and when a person suffers from aphasia, it is in this part where a lesion is regularly found in a postmortem examination. It is clear, then, that the growth of speech during the second year of life is coincident with and dependent on the inner development of this part of the brain; and since this part of the brain is also closely connected with the hands, but more closely with the right than with the left hand, it is to be expected that speech functions bring about activities of the right hand. Thus we understand why certain movements of the baby's speech organs are accompanied by pointing movements of the right hand, and why grown people, too, so frequently accompany their talk by gestures of the right hand.
As to the time of the development of the right hemisphere of the brain in comparison with the left, we are entitled to a conclusion from analogy. The human brain with its complex functions is not ully developed until years after birth. The brain of larger animals of a
( 179) longevity comparable to that of man, with its simpler, but no less important functions, is fully developed sonic months after birth. May not a similar rule govern the development of the left and the right hemispheres? The temporal part of the left hemisphere, with its highly complex speech functions, is not fully developed until years after birth—so much we know. By analogy we conclude that the symmetrically corresponding part of the right hemisphere, with its simpler, though no less important functions, develops to maturity at a much earlier period. If this is so, activity of that hand which is governed by the right hemisphere, must become conspicuous at a much earlier period than activity of the other hand. Indeed, the left hand, whose muscles are closely connected with the temporal part of the right hemisphere, is the preferred member in the activities of the first few months after birth. Thus the fact that a normal human child is at first left-handed and then changes into being right-handed, to remain so during his life, appears plain enough.
Movements of the speech organs are likely to be accompanied by pointing movements or other gestures of the right hand, or of both hands because of the nervous co-ordination of the hands, so we said. Some languages, especially the English, habitually put an enormous vigor into the enunciation of one definite sound of each word or sentence. The English language, that is, is a strongly accented language. According to our law of nervous deflection we might expect that the strong nervous flux leading to the enunciation of the accented sound should interfere with the execution of the hand gesture. It seems that this explains the absence, or at least remarkable infrequency, of gesticulation in speakers using the English language. In the French language, on the other
( 180) hand, there is no accent worth mentioning, and the reflex gestures of the speaker are therefore fully preserved. Accent is thus a substitute for gesture. This explanation seems more generally applicable than the customary one referring to racial differences of temperament as the exclusive cause of the difference in question. Such a difference of "temperament" would remain both ethnologically and psychologically rather mysterious.