The Fundamental Laws of Human Behavior
SIXTH LECTURE
Max Meyer
Table of Contents | Next | Previous
Tension of the total muscular system not interfering with special activities. The motor point not identical with the point that is moved. Fewer reflexes, more instincts in higher animals. The compounding of nervous elements into groups, of these groups into larger groups, and soon, into a single nervous system. The nervous system of a worm. Nerve centers. Lower and higher centers. The nervous system of a crayfish. The brains offish, frog, bird, and mammal. The nervous system of man. The cerebral hemispheres of man.
SINCE we have had occasion to mention again at the end of the last lecture the fact that every nervous process permeates, although for the most part very faintly, the whole nervous system and thus, in a manner, reaches every motor point, we may explain briefly, that this does not conflict with our observations of actual animal behavior. A slight degree of tension of all our muscles in no way interferes with specific activities involving a few definite muscles. Indeed in practically all our normal activities we find that at least one group of muscles other than those which actually pull is quite noticeably under tension, namely the so-called antagonistic muscles. Raise your arm in order to take off your hat. It can be easily observed, and there is no doubt whatsoever, that during the upward movement of the arm not only those muscles are under tension which pull the arm towards the head, but also those which under
( 70) other conditions might pull the arm towards the body. The latter muscles, if we put our fingers on them, do not feel relaxed, flabby, but somewhat stiff. Only, the former muscles feel stiffer, they are under greater tension, and the arm, therefore, follows the direction determined by them. Under these circumstances, a general but faint permeation of the whole nervous system by any nervous process, however definite the activity for whose sake this process exists, involves by no means a contradiction.
Speaking of muscles, it may be well to make a remark here which will help to avoid possible misunderstandings later. When a nervous excitation causes a movement 9f a definite part of the body, for example, of the arm, the motor point of the nervous path is not necessarily located in that part of the body, in the arm. Indeed, if a movement of the arm as a whole is in question, the motor point (more correctly, of course, points) must be in the muscles located at the shoulder and the front or back of the chest. The muscles located in the arm can not move the whole arm, but only the lower arm. The muscles located in the lower arm can not move the lower arm, but only the hand and the fingers. These examples may suffice to call attention to the fact, which is self-evident, that in highly developed animals the motor point must not generally be looked for in that part of the body of whose activity the nervous process is said to be the signal.
We have thus far spoken of the fundamental facts of behavior in higher animals under two headings, that of reflex and that of instinct. We call an animal the higher, the fewer simple reflexes it has—relatively—and the more instincts. Since this difference in the relative number of simple and compound reflexes, that is, of reflexes proper and instincts, depends, according to our previous expositions, on definite structural peculiarities of the nervous
(71) system, it seems interesting and promising to compare lower and higher animals with regard to the structure of [heir nervous systems. It is not a priori necessary, but probable, that these structural differences become apparent to the eye of the anatomist and zoologist, and if they do, they permit the correctness of our previous expositions to be tested.
As our lowest type may serve the nervous system of an earth worm. Figure 19 shows the arrangement of the wore bulky masses of the nervous system, omitting the many fibers which are found scattered over the body and which obviously serve as sensory and motor neurons.
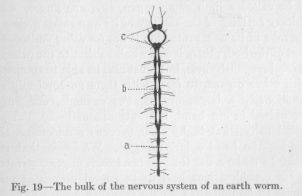
What we see in the figure represents, therefore, in the main, merely the bulk of those connecting neurons which are added to the sensory and motor neurons in order to complete the reflex arches, and of those which serve to establish connections among the reflex arches, in accordance with the principles which we have discussed. We have seen that the frequent necessity of a body acting as a 'whole, the necessity of local responsiveness as found in the reflexes, and the presence of selecting and collecting agents
( 72) called instincts, have as a consequence a structure of the nervous system like the diagrams of Figures 16, 17, and 18. Let us, at present, remember especially Figure 18. We notice there a number of sensory and motor points. Some of the reflex arches are united by the connecting neuron S2abcd M2abcd. The significance of the formation of such a group is obviously, together with the insurance of local responsiveness for each reflex arch, a unification of the group of reflex arches for the purpose of co-operation. It is clear that co-operating reflex arches, as a rule, belong to, and so far as possible are located in, the same part of the body. Any part of the body which deserves to be called a limb or an organ, say, one finger, needs unitary activity of the totality of its own reflex arches for the sake of its own protection if not for other ends. This part of the body, then, is as a whole subject to the principle of local responsiveness, but must also be able to co-operate with other organs, to form with them anatomically and functionally a larger organ. For example, our fingers are united in our hand, not only anatomically, but also functionally, that is, by their nervous connections, thus making the hand a larger organ. We know that it is easier to make a fist than to bend each finger separately. These considerations lead us to expect groups of reflex arches to be united as we see two such groups united in Figure 18, where the central sensory points S2abcd and S2efg and the central motor points M2abcd and M2e fg are connected as if they were peripheral sensory and motor points. The larger organ which is only thus made "an organ," enabled to function as a unit in addition to possessing local responsiveness of its sub-divisions, must again, perhaps, be united with, other organs. That is, the whole mass of nervous conductors, sensory or motor, must again be united with similar masses—we might just
(73) as well say, systems—into a larger "nervous system." Where this building reaches its final culmination, depends exclusively on the anatomical development, the anatomical complication, and, accordingly, the demands of the body for nervous conductors to suit its functional needs. A simple way of representing this compounding of sensory and motor elements into groups, of these groups into larger groups, of these into still larger groups, and so on, finally into a single large system, is shown in Figure 20.
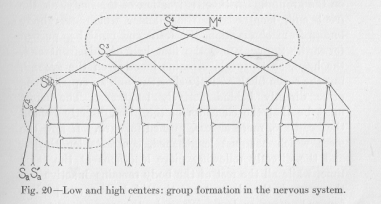
That the total mass of all nervous conductors thus forms a single system, and not two or three or more mutually independent systems, follows from the necessity of each animal body being able to act as a whole. In the figure, each group is composed of only two members, for simplicity's sake. In the body each group must actually contain, of course, a very large number of elements, in accordance with the special anatomical and functional demands. This large and varying number is represented in the diagram almost throughout by two. The dotted lines of the figure will be explained a little later.
( 74)
Let us now make the application to the earth worm. Let us see what kind of a picture we can expect the earth worm's nervous system to present to our eyes. Since the worm's body is long and narrow, we expect that successive pieces, from the front to the rear, should function in relative independence. To make this still clearer, let us remember how the worm moves forward. The abdominal side of the body possesses tiny bristles pointing backwards, so that the body does not easily slide backwards on the ground. If, then, a fraction of the body, at the front end, lengthens in the manner which everybody knows from observation, the front end must be pushed forward. Suppose now the first half of this front end, the head, so to speak, remains inactive on the ground, but the second half actively shortens, and an equally long piece directly behind actively lengthens. The effect must be that the elements of the piece directly behind the head are, more or less, pushed forward. If now the piece which has just lengthened, contracts lengthwise, and the directly following piece lengthens at the same time, while all the rest of the body remains inactive on the ground, the elements of the third piece are pushed forward, more or less. When in the same way every successive piece has been moved forward, we can say that the worm as a whole has made a step forward. It is immediately clear that the nervous system of this animal must be so constructed that the successive pieces can function in relative independence. They must be nervously furnished in such a manner that they can function like so many separate organs, that they possess as wholes what we have called local responsiveness. This means that the reflex arches of each piece must be united into a group, schematically like the groups of reflex arches in Figures 18 and 20. We learn from the figures that this
( 75) unification of the reflex arches adds a considerable number of nervous conductors to those making up each group of reflex arches,—conductors which must be found rather close together, for the sake of the unifying function in question. We must expect, then, to find along the worm, here and there, something like clumps or nodes of nervous substance. Look at Figure 19, where these nodes are apparent. The nervous cord which runs lengthwise through the body shows a number of points where something is added. Where the cord is a plain cord, the addition is simply a swelling, a node, for example, at a. Where the cord is a double cord, in the front part of the body, for example, at b, the addition appears as a kind of tie between the parallel divisions, giving the whole a ladder-like form. These morphological details, of course, do not concern us much at present. The important fact is that we actually find the addition, at fairly regular intervals, of nervous substance to the cord, which confirms our view that the whole nervous cord must be organized in successive groups of reflex arches.
The most conspicuous of these clumps of nervous tissue are the two belonging to the head of the worm, at c. It may be well to mention that the two parallel divisions of the nervous cord are here, between these two nodes, more widely separated, in the shape of an oval, because the alimentary canal, from the mouth, passes through between the two divisions of the nervous cord. If we can speak, in the case of the earth worm, of instincts, that is, if there are nervous functions sufficiently complicated in their sensory and motor aspect and also sufficiently unitary to be called instincts, they must be related to the head rather than to any other part of the body. The head is, even in the earth worm, the most complicated and most important motor organ. Food is taken in by the head.
( 76) The direction of locomotion is determined largely by the direction taken by the head. The head, therefore, must contain many important groups of muscles; but also many important sensory points, for the taking in of food and the direction of locomotion are determined by the nervous stimulations coming from the external objects surrounding the head. It is quite natural that all these important sensory and motor points must be connected by a highly complicated—and therefore somewhat bulky —system of nervous conductors, forming out of the many reflex arches as many different functional groups with manifold interrelations as the life activities of the animal require. This, then, is the cause of the accumulation of nervous tissue in the head, which far surpasses the accumulation of nervous tissue at any of the other nodes of the cord just mentioned. That there are two such accumulations in the head is, from our point of view, a mere accident. It is, of course, the consequence of the anatomical distribution of the head organs. If the anatomy of the head were different, both masses of nervous tissue might appear united into a single node correspondingly larger. And if the anatomy of the head were still different, there might be three, four, or more nodes, each correspondingly smaller.
A name for these nodes has long been adopted, which we must use here too in order to adjust ourselves to the common usage of language. They are called nerve centers. We need not discuss the value of this particular name,—it has become so common that we cannot escape using it. However, it is well to keep any literal meaning of the term entirely out of mind. A "center" has no more literal meaning for us in the nervous system than a "central" has in a telephone system. We know what it is convenient to mean by it, in the one case as in the other. It is cus-
(77) -tomary to speak, not only of nervous centers in general, but of higher and lower nerve centers. The significance of this distinction can now be easily understood. If a "nerve center" is a system of neurons serving the purpose of uniting reflex arches into groups and such groups again into more comprehensive groups, it must be called the higher, the farther removed it is from the sensory and motor points of the body. Still, the word center can be used in a broader or in a narrower sense. In Figure 20, we may call all the neurons enclosed within one of the dotted lines a center. Which of them is higher, more removed from the peripheral sensory and motor points, is immediately clear to the eye. Or, we may take the term center in a narrower sense and include only the connecting neuron between a central sensory point (for example, S2ab ) and its corresponding central motor point (M2ab ), and the adjoining parts of those neurons which from this central sensory and this central motor point run in the peripheral direction. In this sense the neuron S2ab M2ab plus the adjoining parts of the four neurons S1a S2ab , S1b S2ab, M2ab M1a, M2ab M1b would be a nerve center as represented in this diagram. (The reader is requested to supply the lettering of Figure 20 according to the rules given on page 49.) In this narrower sense likewise the distinction between higher and lower nerve centers is immediately clear to the eye. The nerve center of which we just spoke, containing the points S2ab and M2ab, is much lower than the nerve center containing the points S4 and M4. It seems that we may use the term nerve center in either sense without fear that any consequences arise which might make our future discussions ambiguous.
We need not prove that the so-called higher animals possess higher nerve centers than the lower animals, for the greater complication of their nervous system and the
( 78) resulting greater complexity of their life activities is the very reason why certain animals are called higher in comparison with others which then, of course, are called lower animals.. But just on account of this fundamental significance of the terms lower and higher animals, it is exceedingly interesting to study the appearance of the bulk of the nervous system in a series of higher animals as we have studied it in a rather low animal, the earth worm. Let
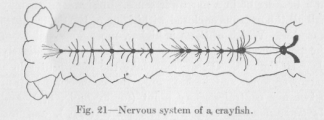
us now use as an example a crayfish (Figure 21), which has a somewhat higher position in the animal scale than a worm. There is no essential difference between the nervous system of the crayfish and that of the earthworm. We notice, however, that the bulky parts of the nervous system, the nerve centers, are relatively bigger. This is to be expected, since the crayfish has numerous appendages, specialized organs which must co-operate for various ends and could not co-operate if their systems of reflex arches were not combined by connecting neurons into a sufficient number of systems of a higher order.
The vertebrates have their more important sensory and motor organs still more concentrated in the head than the articulate and lower animals. The reflex arches of each of these organs must be united into a group, and these groups, again, must enter into manifold combinations in order to serve the more varied needs of a more complex organism. Accordingly we find in the head of a
(79) fish that particularly large accumulation of nerve centers which we call the brain (Figure 22). That this group formation of reflex arches, this development of higher and higher centers is most conspicuous in the head, is the direct result of the greater importance of the sensory and motor organs of the head than of the other parts of the body. The important reflex arches of the head must be well co-ordinated among themselves, but the systems of nervous conductors thus formed must also be well co-ordinated with the less important reflex arches in other parts of the body. However, among the latter reflex arches
there are a certain class which do not require a particularly close connection with the reflex arches of the head. They are the reflex arches which serve the so-called vegetative or visceral functions of the body. Let us make this clear by examples. The approach to a particular article of food is controlled mainly by the reflex arches of the head: it. is a response to sights, sounds, odors received by the sense organs of the head. But the approach could not take place without the co-operation of the reflex arches on which the locomotor organs depend for their function; and these organs are to be found in the remainder of the body rather than in the head. On the other hand, there is scarcely any reason why the intestinal activity of digesting the food should be enhanced or impaired or otherwise influenced during this or any other specially directed
( 80) locomotion of the body, or why this special locomotion should be influenced by the intestinal activity, save the extreme cases of au empty or an already overloaded stomach, to use familiar language. We are not surprised, then, to find in any animal the visceral or interoceptive nervous system rather separated from the locomotor or exteroceptive nervous system, and to find in the nervous accumulation of the head which we call the brain "the center" not so much of the whole nervous system as of the latter part only, of the exteroceptive system.
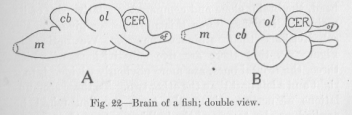
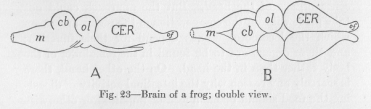
In comparing different vertebrates, let us consider of the whole nervous system this part accumulated in the head, the brain, alone and compare the relative size of its main subdivisions. For this purpose we give to the sub-divisions their ordinary names. We notice, in Figure 22, that there are five sub-divisions, of which three, the frontal ones, are more obviously divided into a right and a left part than the other two. To these accumulations of nervous tissue the term ganglion might be applied, as also to any of the bulkier parts of the nervous systems of the crayfish and the earth worm in Figures 21 and 19. Or they may be called lobes, a tern frequently applied to sub-divisions of the brain. Thus, in Figure 22, of means "optical lobe," of "olfactory lobe." In the same figure m stands for the "medulla," joining the cord, also called bulb because of its shape, cb for "cerebellum" or small brain, CER, for "cerebrum" or large brain.
( 81)
Compare with these ganglions of the fish the same ganglions of the frog, as shown in Figure 23. Their relative size has changed in favor of one, the cerebrum. This is still more obvious in a still higher animal, a bird.
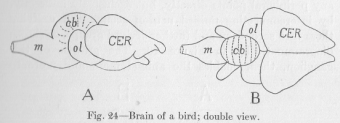
The two halves of the cerebrum, the so-called hemispheres, arc now, especially in the view of Figure 24, B, the most conspicuous part of the whole. The same development continues, when we pass, in Figure 25, to a mammal. The hemispheres of the cerebrum begin to look as if they were the whole brain. The optical lobes have indeed
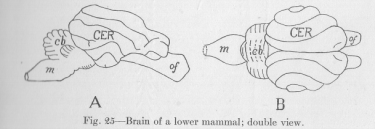
been so completely overlapped by the ever growing hemispheres that they have disappeared from sight. This continued growth of the same single ganglion—quite aside from a continuous, but less marked growth of all of hers—through the various stages of evolution of the vertebrates illustrates a principle different from that
( 82) which requires a bulkier nervous system for an animal possessing a greater number of sensory and motor points. This continued growth of a single ganglion can have a meaning only if the ganglion thus growing does not serve any peripheral points directly, but exclusively indirectly, by interconnecting unified neuron groups and unifying then into secondary and further derived groups, as shown diagrammatically in Figure 20. The growth of this ganglion, then, enables the animal more and more to
( 83) react at any motor point to an excitation occurring at any sensory point whatsoever, without losing its indispensable local responsiveness.
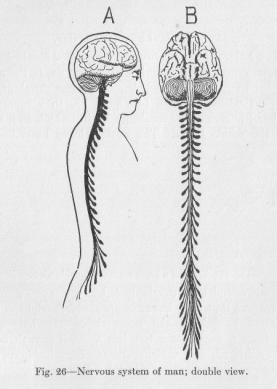
If we compare (Figure 26) the bulky part of the nervous system of man, that is, the part which can be cut out of thebody with comparative ease, and the bulky part of the nervous system of a worm (Figure 19), we see that they are not unlike in appearance except for the fact that to a nervous system like that of the worm man has added the enormous mass of nervous tissue of the cerebral hemispheres (and, we might add, to a lesser extent the cerebellum) serving no other purpose than that explained iii the last paragraph. Along the two sides of the spinal cord, in Figure 26, there are two rows of ganglions somewhat separated from the cord. They obviously group the reflex arches of each side and thus make the organs on one side of the body relatively independent of each other and of those of the other side. Many of our activities, it is true, are of the symmetrical kind, requiring co-operation of the motor organs symmetrically situated. Rut not a few of our activities are one-sided. The reflex arches of a certain region of either side are therefore united iii a ganglion before coming into connection with the nervous conductors of the spinal cord.
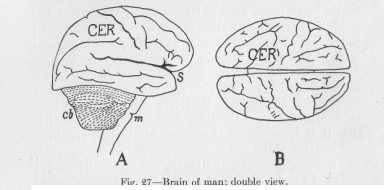
Figure 27 shows the enormous development of the cerebral hemispheres in man as compared with those of the lower vertebrates represented in Figures 22, 23, 24 and 25. The cerebrum has grown to such an extent that it . hides practically all the other parts of the brain, so conspicuous in the fish, the frog, and the bird. The cerebral hemispheres, which in the mammals have fallen sideways over the other original ganglions of the brain, have further grown especially in the forward direction, thus covering their own lateral parts and forming the
( 84) large Sylvian fissure (at S in Figure 27). Only toward the back a piece of the cerebellum is still left uncovered. The cerebrum has become practically the whole brain. Its growth, taking place in the brain exactly there where the most pronounced growth took place during the immediately preceding period of evolution, has every time served to make possible interconnection of the highest centers by still higher centers, thus bringing about an ever increasing possibility of any imaginable group of the seemingly most unrelated motor points being functionally
governed by any group of seemingly most unrelated sensory points. We need not suppose, however, that in this process of complication identical with unification, evolution has reached its highest possible mark. We should over-estimate our race if we thought so. There are many indications—some of them we shall have to mention later that the functions of innumerable reflex arches and groups of reflex arches are even in the human race still practically independent of, never governed by, the function of those, belonging to the majority, which are already effectively unified by their connections with the brain and especially with the cerebral hemispheres.