The Fundamental Laws of Human Behavior
SECOND LECTURE
Max Meyer
Table of Contents | Next | Previous
Contraction at a point other than that of stimulation. The moth and the light. A steady stimulus causing a periodic motion. The need of a nervous system proportional to the development of special motor organs. Sensory and motor points of the body. Nervous connections represented graphically by arches. Cells the biological elements of structure. A nerve cell not a cell but a part o f a cell. Neuron, fiber, ganglion cell. Types of neurons. Collaterals. Terminal arborization and dendrites. White and gray matter. Relative unimportance of the ganglion cell for a theory of mental life.
LET us consider, quite apart from any special problem, in what manner and for what purpose strings conducting like telephone wires could be serviceable in an animal's body. It is plain from the foregoing that they are needed only in case the contraction is to occur not at all at the point of stimulation, but at soma other point. This can be brought about only by conducting away the excitation from the point of stimulation by string-like tissues which cannot themselves contract, but possess a greater conductivity than ordinary, undifferentiated tissues. Carried to an other point, the excitation can there cause the contraction desired. This kind of function is necessary in all the more highly organized animals. Take an insect, a moth for example. We know that the most striking behavior
( 17) of a moth is its flying towards any source of light. This is the result of the nervous connections between I he wings and the eyes. The right eye is connected (if not exclusively, at least better) by nervous strings with the muscles of the left wing, the left eye with the muscles of the right wing. If the moth has the source of light on its right side, the right eye receives more light and consequently a stronger excitation than the left eye. The left wing then beats the air more forcefully than the right wing, and the axis of the animal is turned to the right until both eyes are excited by the light with equal intensity, that is, until the moth flies directly towards the light.
We need not discuss here the question as to the value of this instinct (instinct is the name given to such connections between sense organs and motor organs) to the moth. We may take this value for granted in spite of the fact that millions of moths are destroyed because of this instinct of flying toward the light. Sources of light destructive to moths on the surface of the earth are a vary recant invention of mankind, for which nature cannot be expected to have made provision in giving the moth its biological inheritance. It is not difficult to imagine that this flying in the direction of more strongly illuminated or lighter objects rather than of darker ones aids the moth in obtaining food. So much is plain I hat it could do a moth, whose anatomy is (relatively) so highly developed, no good whatsoever if an excitation caused by light in the region of the head would cause a contraction of the tissues located in the same place. In order to be of any value to the animal it is necessary that I ha chief sensory areas, the eyes, and the chief motor organs, the wings, be connected with each other by differentiated tissues of the conducting kind, by nerves.
( 18)
It may not be amiss, even at the danger of getting a little side-tracked, to discuss another fact which is of great significance for the behavior of animals. We said that the movement of the moth's wings was caused by the excitation occurring in the eye. The question may be asked: How can a rhythmical movement like that of the flapping wings be caused by a continuous excitation like that in the moth's eye? We have no need to explain this here in detail, but it is important to point out, that such a transformation of something continuous into something discontinuous is an exceedingly common occurrence in nature. It is especially important to note that it occurs in the inorganic world, the dead part of nature, as frequently as in the organic world, in living nature, so that we cannot be accused of having neglected the possible claims for recognition of any so-called vital or mental forces when we simply stated that the wings flapped merely because of light falling steadily on the animal's eye. Let us take from the inorganic world a few examples. The wind passing steadily over the surface of the ocean does not cause, by friction, simply a motion of the surface water in the same direction and a compensatory movement of lower layers of water in the opposite direction. It causes, as we all know, a motion of the particles of water which takes place, only to a slight extent in the horizontal direction of the wind, mostly in a vertical direction, up and down, causing waves which periodically rise and fall a considerable height. Or, when we blow a whistle steadily, the result is a rhythmical movement of the particles of air enclosed in the whistle, a physical sound. When water flows very slowly from the faucet in our kitchen, it does not fall in a continuous and very narrow stream, but in periodical drops. Air blown under water through a tube, similarly rises in periodical bubbles.
( 19)
Nobody thinks that such a transformation in these cases requires any hypothetical vital or mental forces. To assume any such forces in the case of muscular activity is equally unnecessary. What we have said about nervous excitation in the eye causing rhythmical motion of the wings is really all that need be said, unless we are specially iii (crested in the details of physiological science.
Let us return to our problem as to the kind of a nervous system which could be regarded as an acceptable gift by our snail or any other animal. We saw that only one kind of behavior is impossible to the snail without a nervous system, namely, a contraction at one point of the body in response to an excitation started at a different point, without any contraction occurring at this latter point, or at least, without a contraction occurring at the point of stimulation with any force approaching that of the contraction at the former point. For example, if the tip of one of the tentacles of a snail—let us think of a snail with tentacles—is affected by a certain stimulus, say fire, it might be safer for the animal to respond by a strong backward movement of its locomotor organs, however far these are from the point of stimulation, than to respond strongly by a contraction of the stimulated tentacle and weakly by action of the locomotor organs. We see at once the close connection between the existence of a nervous system and of highly developed special organs, especially of locomotor organs. Higher animals, having legs, must indeed, because they have these special organs, respond to stimuli occurring at certain excitable points of the body by a forward movement of the legs, to stimuli at other excitable points by a backward movement of the legs, "and by no other motor reaction. The snail, which has scarcely any specialized motor organs, just on this account does not absolutely need
(20) a nervous system. So much nervous tissue as the snail possesses, serves minor purposes which do not much concern us here.
On the other hand, if an animal has specialized locomotor and other motor organs, fins, wings, or legs, with double sets of muscles, for forward and backward motion, its nervous system must be designed according to the following plan and can not be designed in any other way without defeating its purpose. Certain excitable points of the body must be connected by conducting strings with certain contractile tissues located in definite points of the body; other excitable points must be connected with certain other contractile tissues of the body. If we simplify our way of expressing this, we may say: Each sensory (that is, excitable) point of the body must be connected by a conducting string with a definite motor (that is, contractile) point of the body. Let us remember, however, that the facts are riot quite so simple as they are expressed in these words. Actually, a single sensory point is scarcely ever excited in isolation, and a single contractile point, a single muscle fiber, never contracts while all other fibers remain at rest. However, general statements of fundamental facts for the purpose of remembering and reflecting upon them in the abstract, are always artificially simplified. Otherwise they would be of little value to our thought, which is limited in capacity. All scientific laws, even the greatest and most famous of them, are artificial simplifications. This justifies, then, our speaking of the connection of one sensory point with one motor point as if such a simple nervous connection were possible.
We may represent such nervous connections graphically as in Figure 5. Each sensory point S is connected with a motor point M by a conductor, represented in the figure,
( 21) of course, by a line. That this line has the form of a lint arch is not essential. This special form of the diagram has been chosen here because of its convenience, which will later become even more evident. Since there are a
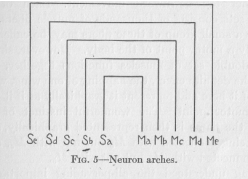
great many sensory and motor points, the diagram ought to contain a great many arches. In our figure they are all represented by only five. They are drawn one above the other because this is the most convenient form of drawing them, not because any such connections are likely to be actually found in a similar parallel arrangement. The difference in size of the arches is also a mere matter of convenience, without any actual meaning. If no other assumptions are made at any later time, we shall always regard the arches as representing nervous connections of the same length.
Let us at once apply this diagram. We spoke above of the instinct of a moth of flying toward the light. Take Sb, to be the right eye and , the left eye. Of course, an eye is not a single sensory point, but since it functions in this case as a unit, there can be no objection to speaking of it, and representing it graphically as if it were a single point. Take Mb to be the left wing's muscles and Mc,
( 22) the right wing's muscles. Again we understand that the muscles of a wing are not a single motor point. The muscles of one wing are, indeed, two sets of muscles, so-called antagonistic sets, of which one serves the upward, the other the downward movement. Each of these muscle sets consists of thousands of muscle fibers. One fiber or a small group of these fibers may be called quite correctly a motor point of the body. However, since the whole double set of muscles functions together, brings about the definite effect, the flapping of one wing, we may speak of it here and represent it graphically as if it were a single motor point. The wonderful instinct of flying toward the flame is then represented graphically, simply by Sb being connected with Mb but not with Mc, and Sc being connected with Mc, but not with Mb.
Having understood the all-pervading importance of these conducting strings for the behavior of all more highly organized animals, it is natural to seek for some more detailed knowledge about their growth, their structure, and their function.
The smallest structural elements of which both animal and vegetable organisms consist have for about a century been called "cells." This means literally boxes—we have a box under our house which we call a cellar. The name appears less strange to us on knowing that those structural elements which were first discovered by means of the microscope happened to look like little boxes. These were plant cells. It was, of course, soon found that not all vegetable elements of structure are box-like. Some, for example the long and thin flax fibers used for the manufacture of linen, do not resemble a box. But the name cell had already been adopted by the biologists as a general name for elements of structure and was now applied also to those elements to which it was not applicable in
(23) its literal meaning. It was equally applied to the elements of structure in the vegetable and animal kingdom, and the whole living world was—and is—said by the biologists to consist of cells. Accordingly the strings, which serve as conductors for excitations in the bodies of higher animals, ought to be called cells,—for the sake of distinguishing them from other kinds of cells, perhaps nerve cells. Such, however, is not the case. The term nerve cell has come to mean, unfortunately, something different. We shall at once see what and why.
In its most undeveloped form an individual unit of nervous tissue is a small, almost spherical body (Figure 6,
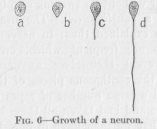
a). As this body grows it becomes pointed in one or more places and sends out a string—like prolongation, which continues to increase in length (Figure 6, b, c, and d), so that it may become easily a hundred thousand times as long as it is thick, reaching a total length of several feet, whereas its thickness is always microscopical. The original little ball from which the string grew out, continues then to exist as a relatively thick swelling of the string. We must remember, however, that it only looks thus, that it did not originate as a swelling of the string. Being relatively bulky, it is not difficult to understand that this thickened part of the string should have attracted the interest of investigators before the exceedingly fine
(24) string. When it was first the object of biological research, its belonging as a part to the long and fine fiber was overlooked. It was studied as an individual thing, and the name cell, generally applied to the elements of biological structure, was applied, instead of to the whole fiber with its swelling, to the swelling alone, which was called a nerve cell. So the inconsistent use of the word cell in its application to nervous tissue, referred to above, came about and is still almost universal.
In more recent years a new, unambiguous terminology has been adopted, which we shall use in the following. We shall call the whole structure, the fiber with its swelling, a neuron, the fiber without its swelling simply fiber, and the swelling alone a ganglion cell. The use of the word ganglion cell is explained thus: In nervous tissues gray looking masses are frequent which, on microscopical examination, reveal themselves as accumulations of swellings with the contiguous pieces of their fibers. It is as if we had a large number of ropes each having a knot somewhere and had taken all these knots in one of our hands. What we then have in our hand might be compared with the accumulation of swellings just mentioned. Such a mass of nervous tissue has long been called a ganglion. Now, it is a peculiar biological fact that these swellings of neurons are not found simply here and there in isolation, but that they are always found in groups, sometimes not very large, sometimes very bulky, —these very ganglions. Since the swellings of the neurons are always found in ganglions, they have been given the name of ganglion cells, which we shall adopt here.
Many are the forms in which the neurons present themselves. Figure 7 shows an assortment of them. The swelling may be at one of the ends as in the case of a and c of the figure, or away from either end as in the
( 25) case of b, d, and e. The long fiber may split into two fibers as in c, or even into more. The swelling may happen to occur just at the point of the division of the
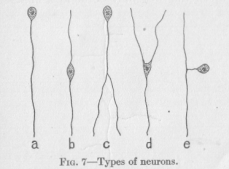
string. In this case the neuron looks like d. The string may in its course turn sideways, form a kind of loop, and continue from the turning point in the original direction. If now the swelling happens to be at the place of the loop, the neuron must look like e. In all these varieties of form we find the same structure, a string with a swelling. Some years ago, when the interest of the histologists was still in the main restricted to the ganglion cell, various kinds of such cells were distinguished according to the number of long fibers which they sent out, and called unipolar (a), bipolar (b) and multipolar (d) cells. Since the ganglion cell has ceased to be regarded as an element of structure in the former sense, these distinctions and names have practically lost their significance. The neuron is essentially a string capable of conducting an excitation from one end to the other. All structural and functional properties are necessarily subservient to this end of conduction.
(26)
Certain features of the neurons, which are not shown in Figure 7, should still be mentioned. We said that the long fibers sometimes split into two fibers. Another breaking up of the fiber may occur in a manner similar
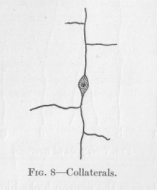
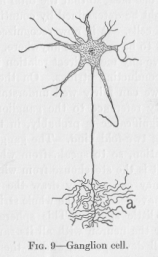
to the way in which a river takes up large tributaries. Long fibers may enter the main fiber as in Figure 8. The tributaries, which most commonly form with the main fiber angles of approximately ninety degrees, are called collaterals. But still another feature of the neurons is to be mentioned. Each ending of a nervous string looks somewhat like the frayed-out end of a thread. The end breaks up into a large number of relatively short branches, the so-called terminal arborization (Fig. 9, at a). In case the swelling of the neuron happens to be located at one of the ends of a neuron, these small branches must naturally come out of the swelling itself. The neuron then looks like Figure 9. The branches proceeding from the swelling are called dendrites, which is a Greek name meaning about the same as the Latin name terminal arborization, namely tree-like branchings. In Figure 9 a neuron is represented whose main fiber is relatively
( 27) short, almost shorter than the dendrites. This shortness, however, is not the rule, but rather the exception; the main fiber, often also called the axis cylinder, usually greatly exceeds the dendrites in length.
There is frequently a difference in coloring between the parts of a neuron. The ganglion cell looks dark, the
fibers lighter. This has given rise to the distinction of white and gray matter in the brain—gray matter taking its name from the presence of numerous dark ganglion cells among the fibers. The popular view, however, of the greater importance of the gray matter than of the white matter is a superstition.
The ganglion cells have a delicate interior structure, and even the fibers are not simple, but possess an interior structure, so that they may be said to consist of fibrils. About the functional significance of these inner divisions
( 28) of a neuron too little is at present definitely known. The question as to the function of the ganglion cell and the fibrous parts of the neuron must be answered at present in a manner very different from that which was customary fifty years ago. It was then often asserted that the ganglion cells were the residences of ideas, each little box the seat of one idea, so that the total mental capacity of a person might be determined by counting the number of his ganglion cells. It is now recognized that an idea can not be said to have its seat anywhere. The ganglion cells do not have any more direct relation to our mental life than the conducting strings. On the contrary, we shall see that we can fairly well understand our mental life without any reference to the ganglion cells. Their physiological significance is probably, in the main, only of the following two-fold kind. The ganglion cell is the point of vegetation, so to speak, from which all growth proceeds, and it is the storehouse from which the neuron in any emergency can quickly draw the means of subsistence. We have seen that the whole string of a neuron grows from a little sphere. This sphere continues to exist even after the neuron with all its ramifications has obtained its full development, and is then the ganglion cell of the neuron. If growth is necessary later, because a branch of the neuron has been cut off or otherwise destroyed, new growth proceeds from that point of the string which is farthest from, but still connected with the ganglion cell. On the other hand, if a conducting string is continually used for hours, changes in the appearance of its ganglion cell occur which probably indicate changes of a chemical nature, called by the physiologists signs of fatigue. It seems that the string, in order to serve continuously for a long time as conductor of an excitation, needs to be resupplied with certain chemicals, and that
( 29) these chemicals are kept in store for the string within the ganglion cell, which, because of its size, is less quickly exhausted than the string. Whether the ganglion cell has any significance in addition to those functions just mentioned, seems doubtful.