Psychology
Chapter 2: The Psychophysical Organism
and the Nervous System
James Rowland Angell
Table of Contents | Next | Previous
The Union of the Psychical and the Physical in the Organism. -- We shall now examine some of the evidence confirmatory of our assertion in the last chapter, that conscious processes and physiological processes are intimately connected in the organism. We shall in this way discover some of the reasons why it is desirable for us at the outset of our study of mental life to learn something about the nervous system, to which subject we shall then devote the remaining portion of the chapter.
Common observation informs us of at least two fundamental types of fact concerning these mind-body relations. We know in this manner (1) that our consciousness or knowledge of the world about us depends primarily upon the use of our senses. A person born blind and deaf has neither visual nor auditory sensations or ideas, and never can have so long as he remains destitute of eyes and ears. By means of the other senses he may be taught much about colours and sounds, as Helen Keller has been; but he never can have the experience which you or I have, when we see a colour or hear a sound, or when we permit a ' melody "to run through our 'heads," as we say, or when we call into our minds the appearance of a friend's face. Indeed, if a child becomes blind before he is five years old he commonly loses all his visual ideas and memories just as completely as though he had been born blind. There is every reason to believe that if we were deprived of all our senses from birth, we could never possess knowledge of any kind. The senses thus hold the keys which unlock the doors of intelligence to the mind, and the
(12) senses are physical, not mental, things. Apparently, therefore, the most simple and fundamental operations of consciousness are bound -up with the existence and activity of certain bodily organs.
Common observation also informs us (2) that the expressions of mind ordinarily take the form of muscular movements which we call acts. We hear a bell and our consciousness of the sound results in our going to open the door. We consider a course of action, and the outcome of our deliberation issues in the form of words or deeds, all of which consist primarily in muscular movements. Strange as it may appear, even keeping still involves muscular activity. It would accordingly seem as though the mind were hemmed in between the sense organs on the one hand and the muscles on the other. If would be a truer expression of the facts, however, to say that these are the tools with which the mind works. Through the sense organs it receives its raw material, and by its own operations this material is worked up and organised into the coherent product which we call intelligence. This intelligence is then made effective in practical ways through the rationally controlled action of the voluntary muscles.
There are other facts of a well-known kind whose precise purport is, perhaps, less evident, but whose general implication of intimate connections between mind and body is identical with that of the considerations which we have just mentioned. We know, for example, that blows and wounds may seriously disturb consciousness, or even destroy it. The similar effects of many drugs, such as alcohol, ether, and hashish, are matters of common knowledge. Even coffee and tea exercise a mild influence upon our psychical mood, and the change in general disposition which frequently follows indulgence in a satisfactory meal is a phenomenon familiar to every family circle. Bodily disease often produces a most marked effect upon the mind, and conversely
(13) different effect upon disease, of a cheerful or a depressed attitude, is a subject of frequent remark.
When we examine the less familiar evidence offered us by certain branches of modern science, we find our previous impressions strongly confirmed. Thus we learn from pathology, the science of disease, that disordered conditions of particular portions of the brain tissue are accompanied by disturbances of definite kinds in consciousness. In this way we learn, for example, that the destruction or disintegration of the tissue of one region in the brain is followed by the loss of one's visual memories, so that one cannot recall the appearance of familiar objects. A similar disorder in another region costs one the control of certain muscles in the hand, etc. The science of anatomy is able to demonstrate structural connections of nerves between these diseased parts of the brain and the sense organs and muscles over which consciousness has lost control, thus supporting the implication of the pathological evidence already cited. Experimental physiology shows us, that by stimulating (either mechanically or electrically) certain brain areas in animals, we can produce movements of definite muscles, whereas by extirpating these regions we can at least temporarily cripple the muscles and render the will powerless over them. By similar excisions of other brain areas we can cripple definite sense organs. Thus pathology, anatomy, and physiology all point to the same intimate relation of mind and body and indicate more specifically than the observations of every-day experience could do a fixed and positive relation between definite parts of the nervous system and such special phases of consciousness as the visual, the auditory, etc.
Moreover, comparative anatomy, comparative physiology, and comparative psychology all converge upon another cognate principle, i. e., that the development of consciousness among various genera and species of the organic world has run parallel with the development of the nervous system.
(14) Taking all these considerations into account, the deliverances of common sense as well as the teachings of science, it is easy to understand why the modern psychologist finds it judicious in his study of consciousness to learn all that he can about the nervous system, the sense organs, and the motor mechanism.
The Nervous System. -- It will assist us in gaining a working idea of the nervous system to bear in mind the fact that its fundamental function consists in "the conversion of incoming sensations into outgoing movements of a kind tending to preserve the creature." Creatures destitute of some form of nervous system are practically incapable of prompt and appropriate adaptation to their surroundings. Plants are thus in large measure the passive victims of their environments. Injury to one part commonly produces little or -no immediate effect upon the rest of the plant. But by means of its nervous system every part of an animal organism is brought into vital connection with every other part. Cooperation becomes the controlling principle in the life activities. This cooperation, or coordination, takes the form of movements made in response to sensory stimulations, and the most highly evolved form of nervous system, such as that of the human being, differs from the very -rudimentary forms, like that of the jelly fish, only in the complexity of the devices by which these stimulations and movements are connected. When we are studying the structure of this system, we should, then, always remember this fact about the coordination of sensations and movements, as the clue by which to interpret even its most intricate arrangements.
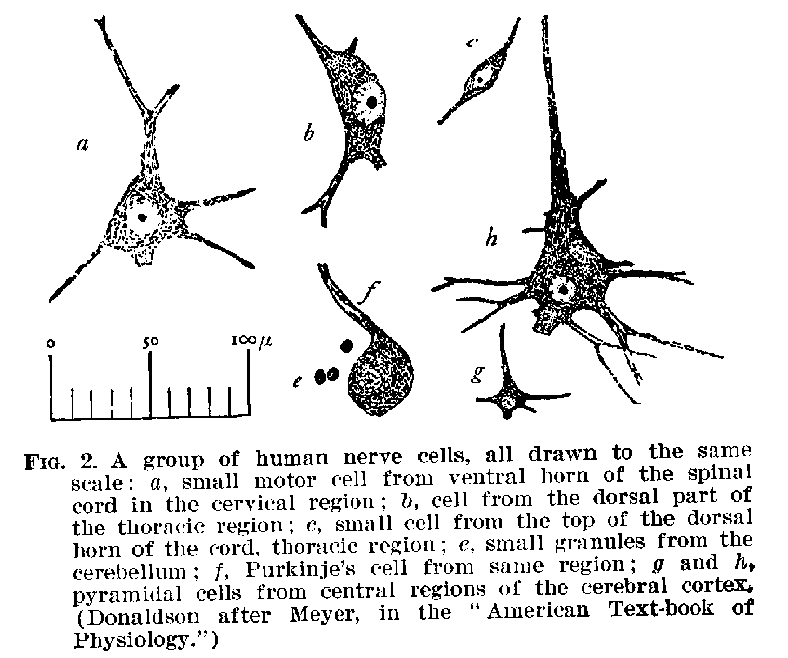
The Elementary Structures.-- The nervous system is made up of nerve cells, with their filamentous elongations which are called fibres. A sketch of certain common forms of nerve cells is shown in figures 1, 2, and 3. It will be seen that they are accumulations of granular protoplasmic masses containing a nucleus, and often within this nucleus smaller
(15) nucleoli, while from their edges are given off filaments of various forms and sizes. These filaments are outgrowths of the cell-body and constitute an organic part of the structure. The whole structure, including both fibre and cell-body, is nowadays called a neurone. The neurone is, therefore, the
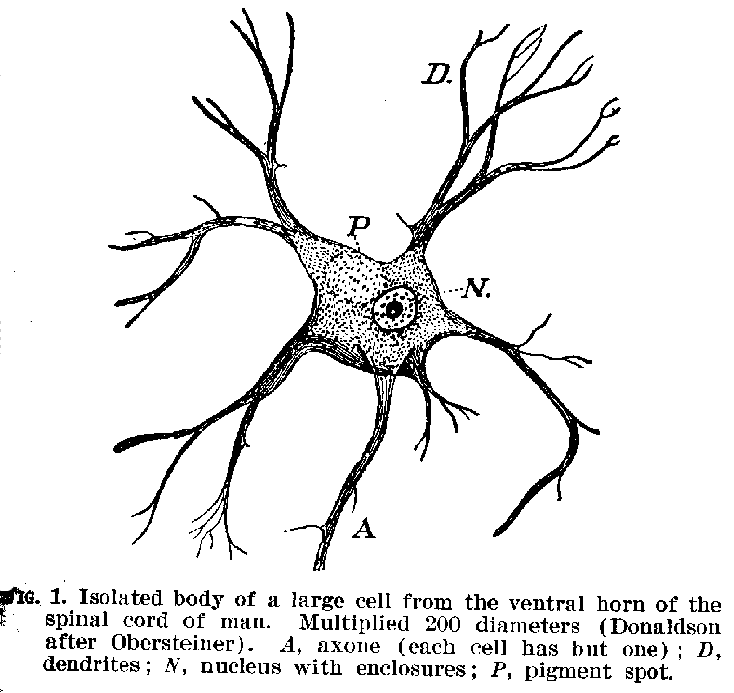
real element of the nervous system. It has been estimated that in the nervous system of the adult human being there are about 11,000 millions of these neurones in various stages of development. Their average volume is probably about .00009 of a cubic millimetre.[1]
(16)
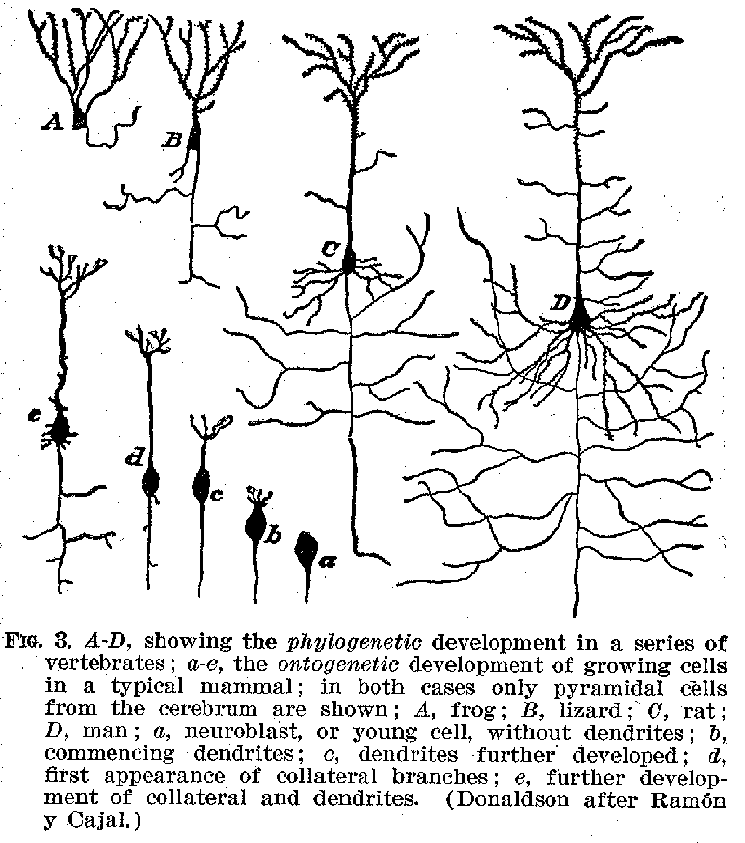
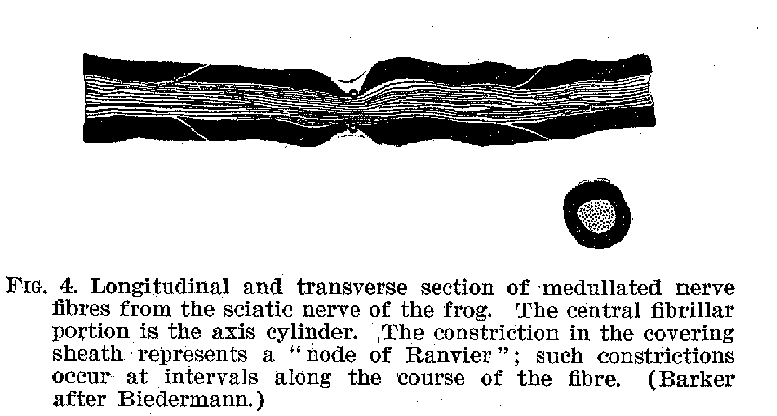
Certain of the fibrous protuberances are called axones or neurites , others are known as dendrites. The axones, as may be seen from figure 3, are generally smooth in their contours, and when they branch, the divisions commonly occur at right angles. Within the central system the dendrites are rougher and branch more gradually from one another, somewhat like the sticks of a fan. The fully developed axones have a peculiar structure, shown in figure 4. The central strand is known as the axis cylinder. This is a transparent mass
which apparently constitutes the true nerve, and conducts nervous impulses from sense organ to nerve centre, and back again from nerve centre to muscle. Outside the axis cylinder is a relatively thick covering known as the medullary sheath. This sheath generally disappears near the cell-bodies and also wherever the fibre terminus approaches other fibre ter
(17) minals. Outside of this again there is a thin membraneous sheath known as the neurilemma.
Although the cell-bodies and fibres are really parts of single organic cells -- the neurones -- their notable difference in appearance is accompanied by a distinct difference in function. Both cell-bodies and fibres are sensitive to stimulation,
are irritable, as the physiologists say, and both possess conductivity. But whereas this exhausts the fundamental functions of the fibres, the cell-bodies are ordinarily supposed to possess the further capacities of reinforcing or inhibiting
(18) the impulses sent to them. Moreover, the cell-bodies seem at times to send out nervous excitation along the fibres automatically, without any detectable external stimulation. It will be seen, therefore, that the cell-bodies are in a sense the
power centres of the nervous system, while the fibres are in the main merely interconnecting mechanisms, putting the several sense organs into relation (1) with the various centres and (2) through these with the muscles.
It is supposed that inside the central nervous system the axones are ordinarily employed to carry impulses away from the cell-bodies, whereas the dendrites probably carry impulses toward them. Outside the central system the afferent fibres leading to the spinal ganglia resemble axones in structure, and so offer apparent exceptions to this rule. In any event the whole nervous system is nothing but an aggregation of neurones with the supporting tissue, called neuroglia, which holds them in place. A nervous impulse originating in the sensory surface of the body, for example in the retina, may be transmitted from one group of neurones to another, until finally it issues, perhaps, from the nerves of the spinal cord, and produces a movement of the foot. This is what would occur if one should step aside upon seeing a heavy object about to fall. In this process of transmitting the
(19) impulse through the nervous system, it is not necessary that the groups of neurones should be actually in contact with one another, although this may occur. But they must at least be close together. Contiguity, if not anatomical continuity of groups of neurones, seems to be a sine qua non of neural conductivity.
The exact physical nature of neural excitement is not known. Various theories have been propounded in the effort to identify it with recognised forms of chemical or electrical activity, but thus far no hypothesis has been suggested which accords satisfactorily with all the facts. Meantime, we speak of the nervous current, the neural disturbance or excitement, in a purely metaphorical way, to cover the facts which we do know, i. e., that physiological activity of a certain kind occurs in the nervous structures, and is transmitted very rapidly from one point to another. In man the rate of this transmission is about 100 feet per second.
 Various Forms of Nervous System.-- When we turn to the
zoologist and the comparative anatomist, we are able to obtain certain interesting facts
about the development of the nervous system throughout the organic kingdom. From such
sources we learn that the simplest types of animal organism, e. g., such protozoans as the
amoeba , possess no nervous system at all. Every part of the surface of the unicellular
amoeba (figure 5) is capable of movement, of assimilating food and excreting the waste
products. This animal's behaviour suggests that other forms of tissue besides nervous
tissue are sensitive and capable of conducting impulses. Undoubtedly this is a fact, and
we must accordingly think of the nerves as simply specialised
Various Forms of Nervous System.-- When we turn to the
zoologist and the comparative anatomist, we are able to obtain certain interesting facts
about the development of the nervous system throughout the organic kingdom. From such
sources we learn that the simplest types of animal organism, e. g., such protozoans as the
amoeba , possess no nervous system at all. Every part of the surface of the unicellular
amoeba (figure 5) is capable of movement, of assimilating food and excreting the waste
products. This animal's behaviour suggests that other forms of tissue besides nervous
tissue are sensitive and capable of conducting impulses. Undoubtedly this is a fact, and
we must accordingly think of the nerves as simply specialised
 (20) forms of protoplasm in which these functions are more highly developed
than elsewhere. In certain of the lower metazoans nerve cells appear with fibres extending
toward the periphery of the body and possessing sensitive terminations. Among the
coelenterates a very simple nervous system comes to light. In bydroids this is merely a
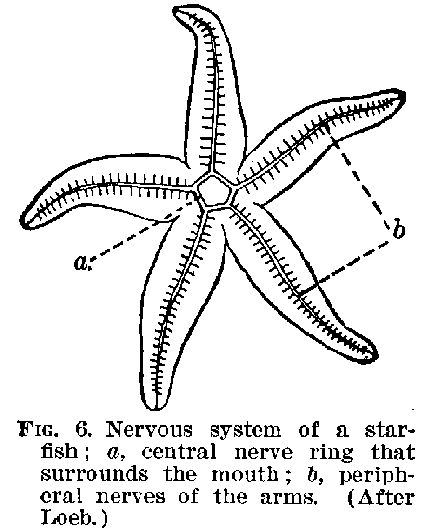
kind of tissue of nerve cells. In echinoderms we meet with a structure like that shown in
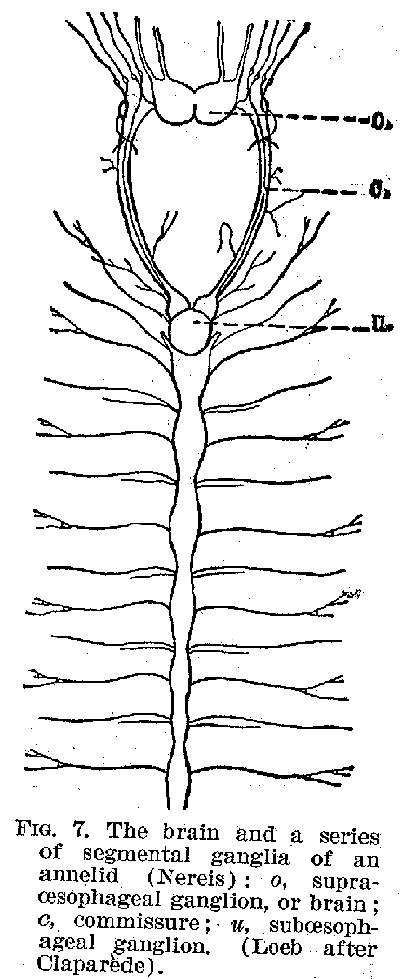
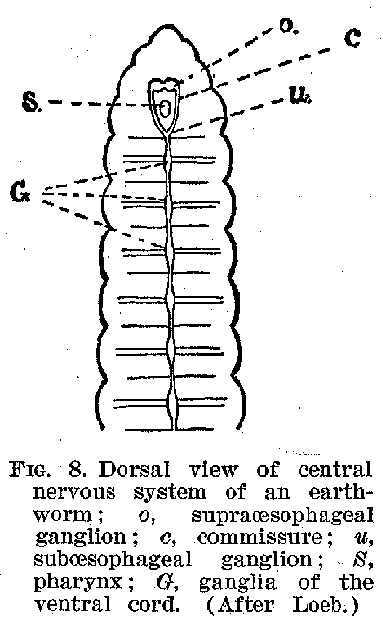
figure 6. But it is not till we get to such forms as the worms that we find a definite
organised centre of control, like the brain or spinal cord. In the annulates of the worm
forms there is not only a centre corresponding to a very rudimentary brain, but also one
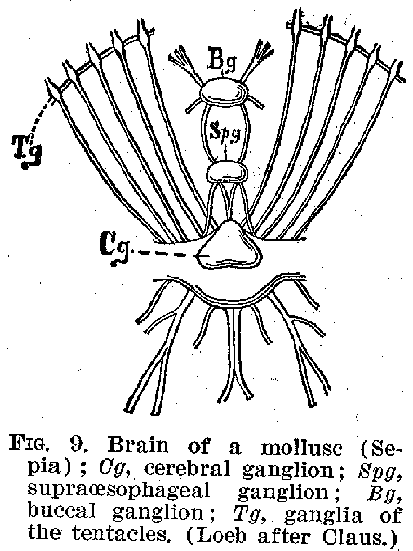
roughly corresponding to the spinal cord. (Figures 7 and 8.) In the molluscs the
development is made more complex by the appearance of these groups of central cells
clustered together in several directions about the brain. (Figure 9.) Even in the lowest
forms of vertebrates, e. g., the acranial amphioxus, we find both a brain and cord.
Passing from the lowest to the highest vertebrates up, for example, through the fishes,
reptiles, and amphibians to the birds and mammals, we meet with every shade of variation
in the development of the several parts of the nervous system. Everywhere, however, from
the most primitive metazoan up to man, the general principle is one and the same
mechanism for connecting sensitive surface organs with muscles.
(20) forms of protoplasm in which these functions are more highly developed
than elsewhere. In certain of the lower metazoans nerve cells appear with fibres extending
toward the periphery of the body and possessing sensitive terminations. Among the
coelenterates a very simple nervous system comes to light. In bydroids this is merely a
kind of tissue of nerve cells. In echinoderms we meet with a structure like that shown in
figure 6. But it is not till we get to such forms as the worms that we find a definite
organised centre of control, like the brain or spinal cord. In the annulates of the worm
forms there is not only a centre corresponding to a very rudimentary brain, but also one
roughly corresponding to the spinal cord. (Figures 7 and 8.) In the molluscs the
development is made more complex by the appearance of these groups of central cells
clustered together in several directions about the brain. (Figure 9.) Even in the lowest
forms of vertebrates, e. g., the acranial amphioxus, we find both a brain and cord.
Passing from the lowest to the highest vertebrates up, for example, through the fishes,
reptiles, and amphibians to the birds and mammals, we meet with every shade of variation
in the development of the several parts of the nervous system. Everywhere, however, from
the most primitive metazoan up to man, the general principle is one and the same
mechanism for connecting sensitive surface organs with muscles.
(21)
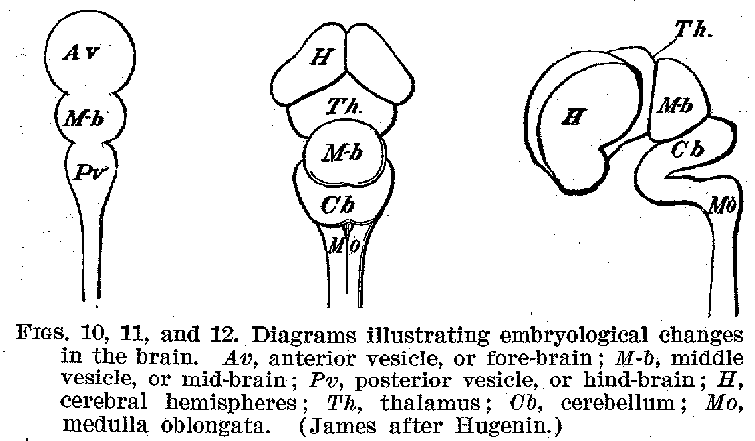
 The Gross Structures of the Human Nervous System. -- If we
were to examine the human nervous system at one period of its development, we should find
it a crude structure of tubular form, with one end enlarged, and slightly constricted at
two zones, as shown in figures 10, 11, and 12. The walls of this enlarged portion thicken
and spread out as they grow, and in one place dwindle away to a mere membrane. In this
manner the various parts of the adult brain are formed, retaining to the end the old
tubular contours. The remnant of the cavities in the embryonic brain and cord become
respectively the ventricles of the developed brain and the canal of the spinal cord. These
cavities remain connected with one another and are filled with the cerebrospinal fluid.
The surfaces of the brain and cord are closely invested with a membrane, the pia mater,
carrying blood-vessels. This membrane is bathed on its outer surfaces by fluids. A tough,
thick membrane, the dura mater, separates the pia water from the bones of the skull
and vertebrae. The portion of the embryonic brain known as the fore-
The Gross Structures of the Human Nervous System. -- If we
were to examine the human nervous system at one period of its development, we should find
it a crude structure of tubular form, with one end enlarged, and slightly constricted at
two zones, as shown in figures 10, 11, and 12. The walls of this enlarged portion thicken
and spread out as they grow, and in one place dwindle away to a mere membrane. In this
manner the various parts of the adult brain are formed, retaining to the end the old
tubular contours. The remnant of the cavities in the embryonic brain and cord become
respectively the ventricles of the developed brain and the canal of the spinal cord. These
cavities remain connected with one another and are filled with the cerebrospinal fluid.
The surfaces of the brain and cord are closely invested with a membrane, the pia mater,
carrying blood-vessels. This membrane is bathed on its outer surfaces by fluids. A tough,
thick membrane, the dura mater, separates the pia water from the bones of the skull
and vertebrae. The portion of the embryonic brain known as the fore-
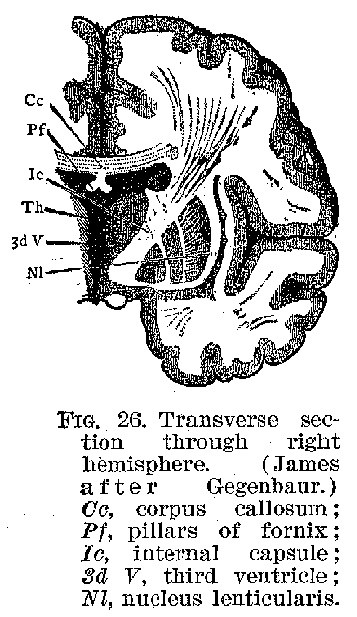
(22) -brain finally develops into the great masses of the cerebrum. The optic thalami, which are large collections of nerve cells with their fibrous connections, also belonged originally to this general region of the brain. The primitive mid-brain changes less in mass during growth than does the fore-brain, and becomes on its under or ventral surface the crura or peduncles of the brain, while on its upper or dorsal surface it C becomes the corpora quadrigemina. The hind-brain develops in its foremost part, dorsally into the cerebellum, and ventrally into the pons. In its lower portions it becomes the medulla oblongata, upon the dorsal surface of which appears the fourth ventricle, with its non-nervous membraneous covering. The spinal cord undergoes the least profound change, as regards its external contours, of any of the embryonic parts of the central system.
When we take the facts of development into account, therefore, it becomes evident that the various portions of the brain, which seem at first glance so hopelessly confused in their relations to one another, are nevertheless all outgrowths of a single relatively simple structure-the tubular embryonic nervous system, whose walls are everywhere made up of neurones and their supporting tissues, the neuroglia. The general form of the brain is complete some time before birth.
The number of neurones, the nervous element, is also com-
(23)-plete at birth. But the maturity of the brain in point of external size is not reached until about seven years of age, and development in the size and interconnections of the neurones goes on indefinitely, certainly with most persons up ito forty years of age.
Following Ebbinghaus and others we may classify the neurones of the central nervous system in three general groups, (1) the peripheral neurones, (2) the subcortical neurones, and (3) the cortical neurones.
 Peripheral Neurones -- The
peripheral neurones, whose cell-bodies lie outside the central system, consist of
sensory cells and their fibrous prolongations of which a part extend inward toward the
centres, and a part outward toward the sensory end-organs, such as the rods and cones of
the retina, the hair cells of the cochlea in the internal ear, the touch corpuscles in the
skin, etc. (See cuts in Chapter V.) The cell-bodies of these neurones are sometimes
situated near the central structures, as in the case of The cells in the ganglia of the
posterior roots of the spinal cord. These cells distribute their fibres to the skin,
muscles, tendons, etc. Sometimes, however, they are in the neighbourhood of the sense
organ, in the case of the auditory nerve, which arises from cell in the internal ear; the
optic nerve, which has its cell-body in the retina, etc. The function of the peripheral
neurones is evidently that of transmitting impulses from the
Peripheral Neurones -- The
peripheral neurones, whose cell-bodies lie outside the central system, consist of
sensory cells and their fibrous prolongations of which a part extend inward toward the
centres, and a part outward toward the sensory end-organs, such as the rods and cones of
the retina, the hair cells of the cochlea in the internal ear, the touch corpuscles in the
skin, etc. (See cuts in Chapter V.) The cell-bodies of these neurones are sometimes
situated near the central structures, as in the case of The cells in the ganglia of the
posterior roots of the spinal cord. These cells distribute their fibres to the skin,
muscles, tendons, etc. Sometimes, however, they are in the neighbourhood of the sense
organ, in the case of the auditory nerve, which arises from cell in the internal ear; the
optic nerve, which has its cell-body in the retina, etc. The function of the peripheral
neurones is evidently that of transmitting impulses from the
(24) sense organs in to the nervous centres, and we need discuss them no further at this point.
Subcortical Neurones. -- The subcortical group involves all the gross structures in the central system lying between the
cortices of the cerebrum and the cerebellum on the one hand and the peripheral neurones on the other. Their function is in general that of furnishing neural mechanisms for reflex acts and for connecting the various parts of the central system with one another. This can be best brought out by examining separately some of the more conspicuous gross structures of this group. After we have done this, we shall turn to the cortical groups, whose functions as general control centres we shall then discuss.
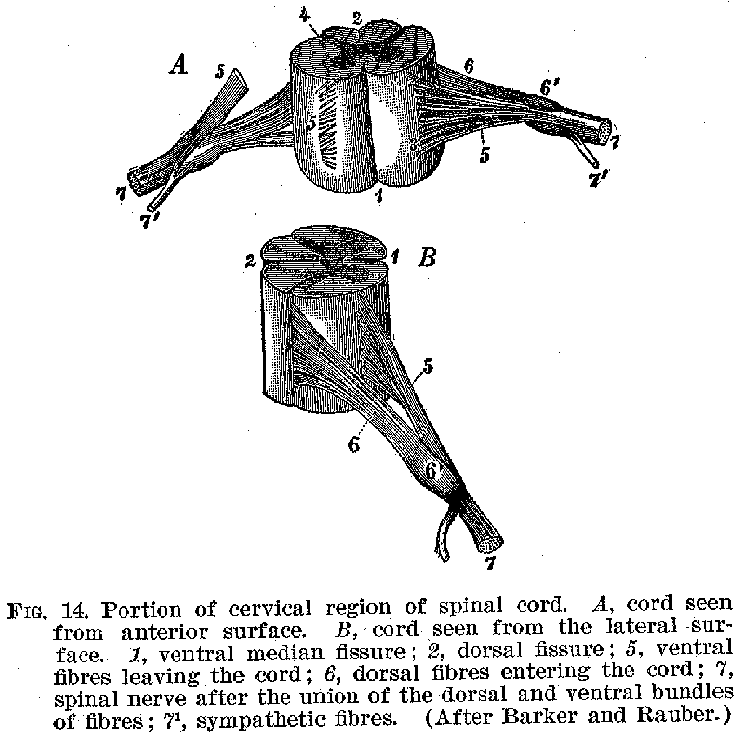
We may first consider the spinal cord (figure 13). If we take a cross section of this organ, cutting through at right angles to its long axis, we find a structure such as is shown in figures 14 and 15. In the central portion, grouped about the spinal canal in the general shape of the letter H, is a great mass of cell-bodies giving a peculiar greyish colour to the region. Outside of this is a thick layer of white nerve fibres. Close examination of the grey matter reveals fibres running out laterally to penetrate the white masses. The
 (25) fibres from the cell-bodies in the ventral or anterior region of the
grey mass pass out from the spinal cord in bundles at the general level of the several
spinal vertebrae. Thence they may be traced principally to the voluntary muscles of the
limbs and trunk. The fibres issuing from the dorsal or posterior region of the grey matter
pass out in similar bundles from the posterior sides of the cord, and thence, after
uniting with the bundles from the motor region, are distributed chiefly to the sense
organs of the skin, joints, muscles, tendons, etc. It may be remarked at this point that
the voluntary muscles, such as control the movements of the hand, are commonly striped
muscles, whereas the involuntary muscles, e.g., those of the alimentary and circulatory
systems, are generally unstriped. The unstriped muscles are mainly connected with the
sympathetic nervous system, of which we shall speak briefly a little later. The striped
muscles contract and relax more rapidly than the unstriped.
(25) fibres from the cell-bodies in the ventral or anterior region of the
grey mass pass out from the spinal cord in bundles at the general level of the several
spinal vertebrae. Thence they may be traced principally to the voluntary muscles of the
limbs and trunk. The fibres issuing from the dorsal or posterior region of the grey matter
pass out in similar bundles from the posterior sides of the cord, and thence, after
uniting with the bundles from the motor region, are distributed chiefly to the sense
organs of the skin, joints, muscles, tendons, etc. It may be remarked at this point that
the voluntary muscles, such as control the movements of the hand, are commonly striped
muscles, whereas the involuntary muscles, e.g., those of the alimentary and circulatory
systems, are generally unstriped. The unstriped muscles are mainly connected with the
sympathetic nervous system, of which we shall speak briefly a little later. The striped
muscles contract and relax more rapidly than the unstriped.
The arrangement of the elements in the spinal cord sug
(26) -gests at once two of its principal functions, and is so typical of the facts generally characterising nervous structure and function that it seems judicious to comment upon it briefly.
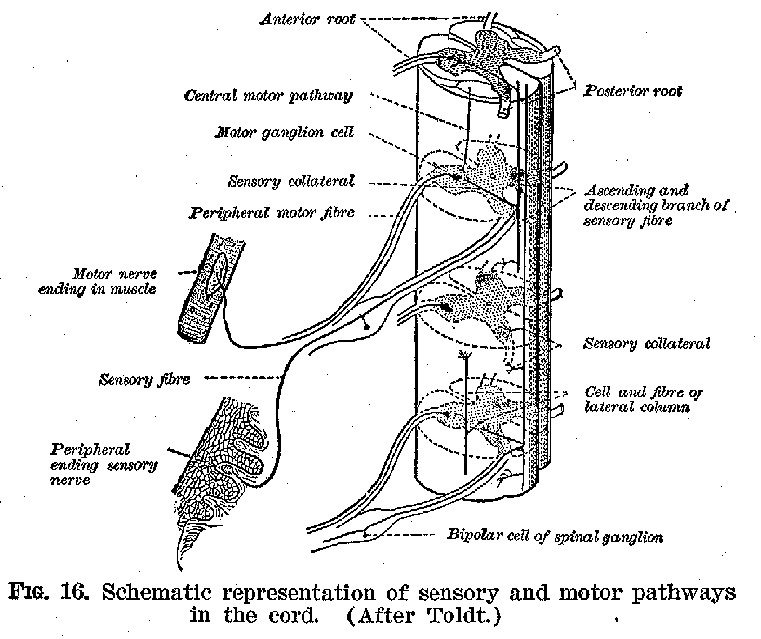
It will be observed in the first place, that in the cord cellbodies, connected through their fibres with the sense organs and the muscles respectively, are in very close proximity to one another. It should be relatively easy, therefore, for an incoming sensory impulse to find its way out over motor nerves and so to produce reflex movements, that is, movements made in immediate response to sensory stimulations, without the guiding action of consciousness. This is precisely what happens, and it is as a reflex mechanism that the spinal cord exercises one of its important functions. As in
(27)-stances of such reflexes we may mention the jerking of the foot when the sole is tickled, and the knee-jerk exhibited when one leg is crossed over the other and the region just below the knee-cap is smartly struck. Furthermore, we should find upon examination that the white fibrous tracts along the external surfaces of the cord connect it with both the higher and the lower parts of the system. (Figures 16 and 17.) It should thus be easy for impulses to pass upward and downward, between the brain on the one hand and the sense organs and muscles on the other. Such ready transmission actually occurs, and it is in this fact that we find the second great function of this organ. The spinal cord is accordingly typical of the central structures in general, in that
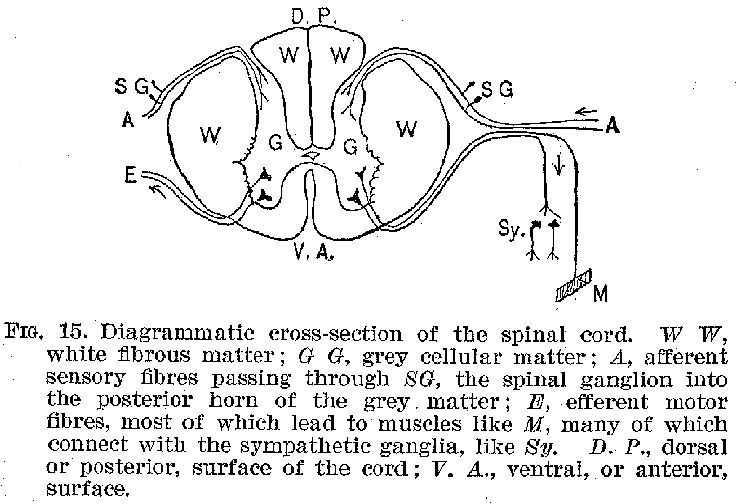
it provides (1) means for the immediate connection of sense organs and muscles and (2) devices for connecting various parts of its own and other nervous structures with one another.
If we were to examine the other subcortical masses lying between the cerebrum and the spinal cord, we should find that, in general, they consist of aggregations of neurones
(28) much like those in the cord, but on the whole less simply and regularly disposed. Thus, the medulla, the corpora, and the thalami all display ganglion groups with sensory and motor
connections. When we come to speak of their specific functions, we are obliged to indulge largely in speculation, because the facts are evidently extremely complex, and our knowledge of the details involved is notoriously incomplete. Moreover, the specialised functions which are sometimes attributed to them in the case of the lower animals are probably in the human being largely usurped by the cerebral cortex. In any event we must always remember that the nervous system is an organic unit, and no part of it ever acts wholly independently of the other parts, nor is any influence exercised upon one part entirely without significance for the other parts. Any mention of specific functions of different regions must always be made with this reservation in mind.
 (29) Thus, the spinal cord undoubtedly contains the neurones whose
innervation is immediately responsible for movements of the hand. But this innervation
itself may originate almost anywhere throughout the rest of the nervous system, so that
any portion of this system may in a particular case contrib ute to the production of the
special motor consequences. To say that a region of the nervous system presides over any
special function is, therefore, simply to say that it is the portion most immediately and
most invariably responsible for it.
(29) Thus, the spinal cord undoubtedly contains the neurones whose
innervation is immediately responsible for movements of the hand. But this innervation
itself may originate almost anywhere throughout the rest of the nervous system, so that
any portion of this system may in a particular case contrib ute to the production of the
special motor consequences. To say that a region of the nervous system presides over any
special function is, therefore, simply to say that it is the portion most immediately and
most invariably responsible for it.
Speaking within such limitations, we may say that there
(30) are two functions of centres lying in the medullic region, about which considerable unanimity of opinion exists. The control of the automatic respiratory movements, and the control of the vaso-motor nerves which govern the calibre of the
arteries, are apparently the immediate possessions of neurones belonging in this neighbourhood. Needless to say,, all these regions are like the spinal cord in containing pathways for
(31) neural excitation to pass upward and downward, between the cerebrum on the one hand and the sense organs and muscles on the other.
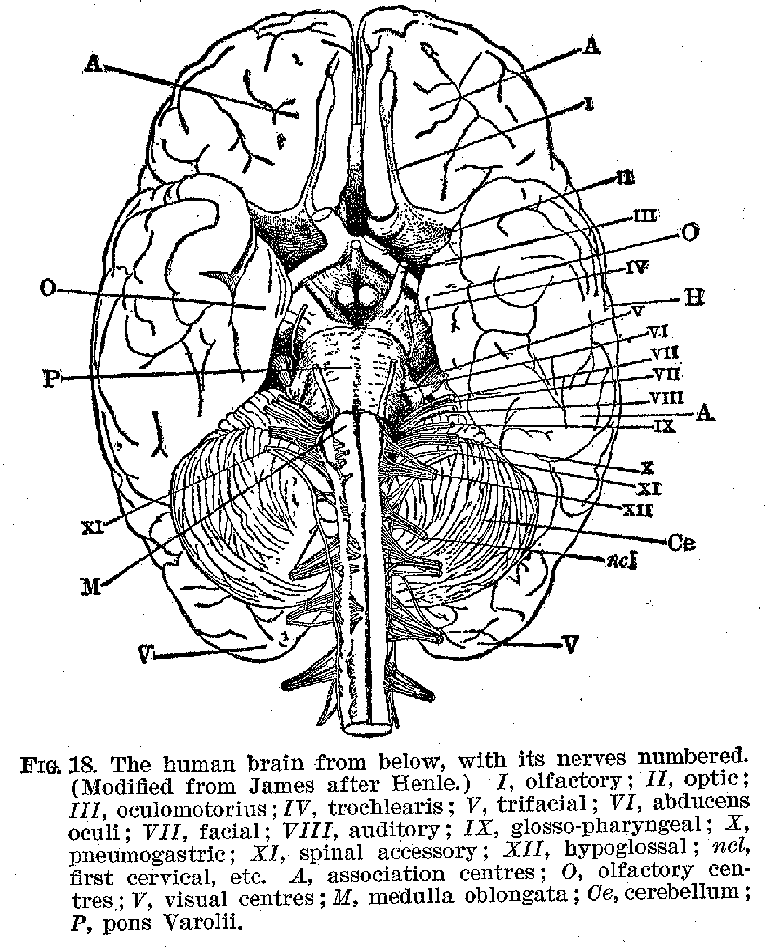
It is in this region of the brain, too, that the cranial nerves are given off, e. g., the nerves of special sense, like the olfactory, the auditory, the optic, the gustatory and trigeminus; such motor nerves as those controlling the eyes, the tongue, the lips, etc. (Figure 18.) It is commonly maintained that the phylogenetic pattern from which the human nervous system has been developed is of a segmental character, each
part of it receiving sensory and motor nerves from relatively distinct regions, or segments, over which they exercise a definite and sometimes exclusive control, In the human being
(32) the motor nerves can, indeed, be classified in this segmental way in accordance with the special muscles which they innervate, e. g., those of the head, the upper trunk, the lower trunk, etc. But the connections of the sensory neurones in man make any such segmental divisions of them very hazardous, so that the application of the segmental idea to the interpretation of our human nervous action is somewhat uncertain.
The Cortical Neurones. -- The Cerebellum. -- So little is known about the operations of the cortex of the cerebellum, that it will not be profitable to discuss it. Suffice it to say that the cerebellum has a very rich connection, by means of both sensory and motor neurones, with the cerebrum and the lower brain centres.
The Cerebrum.-- For the psychologist the cerebral hemispheres are the most interesting and most important portions of the nervous system. From the various lines of evidence mentioned earlier in the chapter, we know that consciousness is connected with this part of the brain in an exceedingly intimate way, and we shall consequently devote some little space to its consideration.
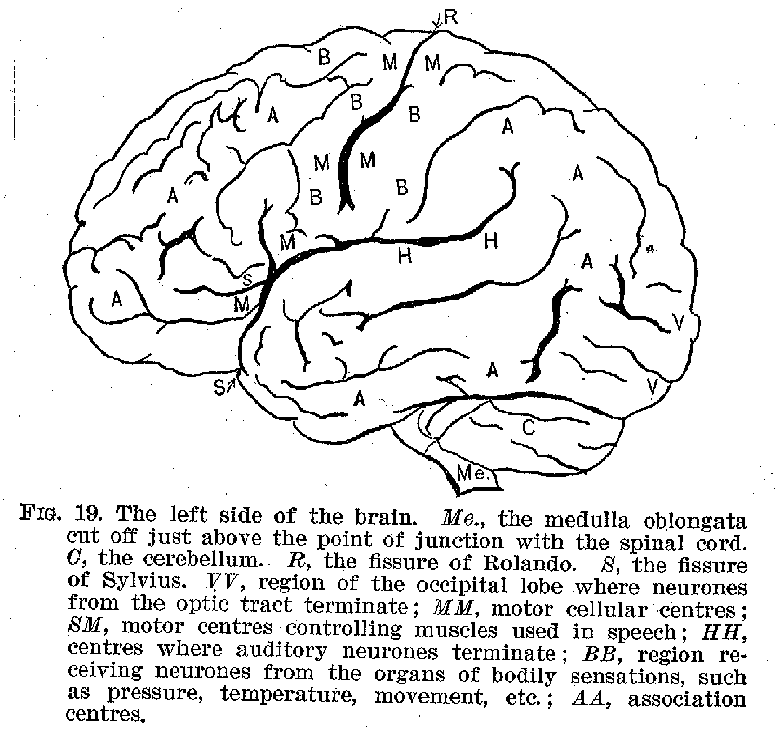
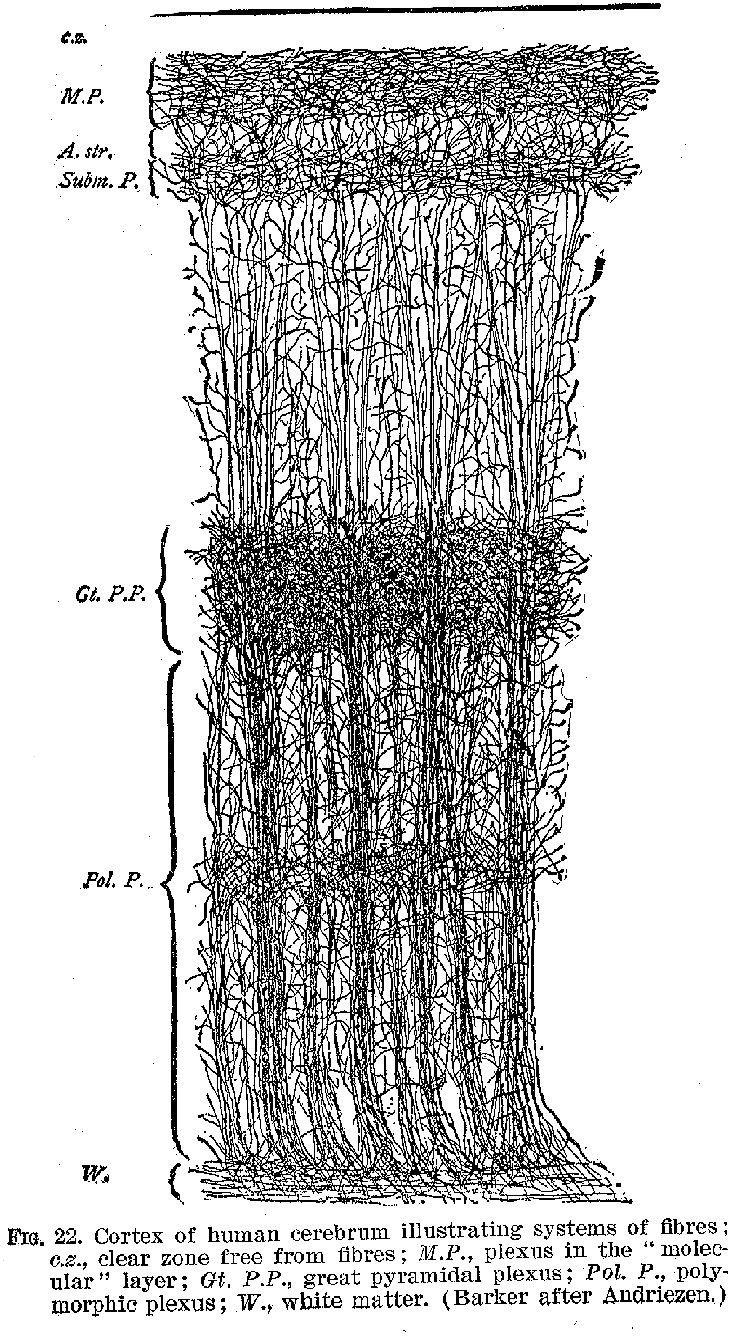
The surface of the hemispheres, called the cortex, and shown in figures 19 to 22, is made up of layers of cell-bodies, with their delicate protoplasmic processes. The extraordinary richness of the dendritic development in the cortical neurones furnishes one of the most marked peculiarities of the human cerebrum, as contrasted with those of animals. This intricate dendritic structure apparently represents the bodily counterpart of those elaborate interrelations among ideational processes, which characterise in general the higher forms of intelligence.
Certain of the cortical areas are known to be in functional connection with sense organs from which they receive stimuli. Thus, the region marked H is in connection with the ear, and receives auditory impressions. (Compare figures 18 to 21.) The region marked V is similarly connected with the retina,
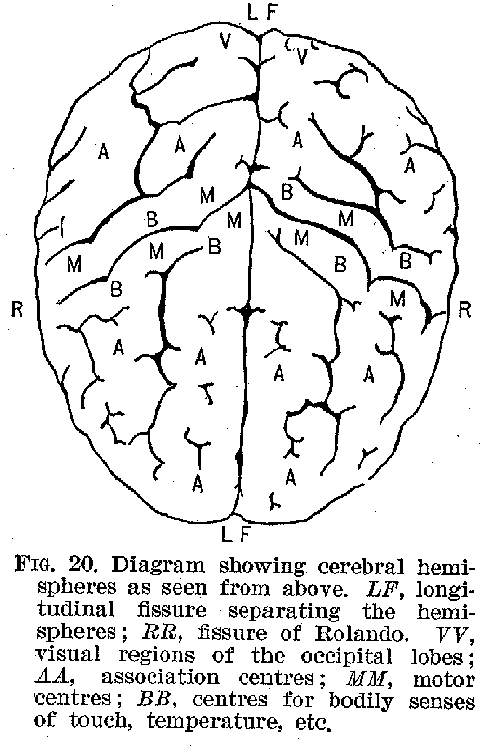 (33) and receives visual impressions. It is reasonably certain that areas
marked O receive olfactory stimuli, while the region marked B is probably that immediately
concerned with the reception of tactual and thermal stimuli. The centres for taste are not
clearly made out. It seems probable, however, that they are in the neighbourhood of the
olfactory terminals. There is reason to believe that ordinarily the peripheral sensory
neurones are in connection with the side of the cortex opposite to that from which they
originate For example, the touch nerves of the left hand find their cortical terminations
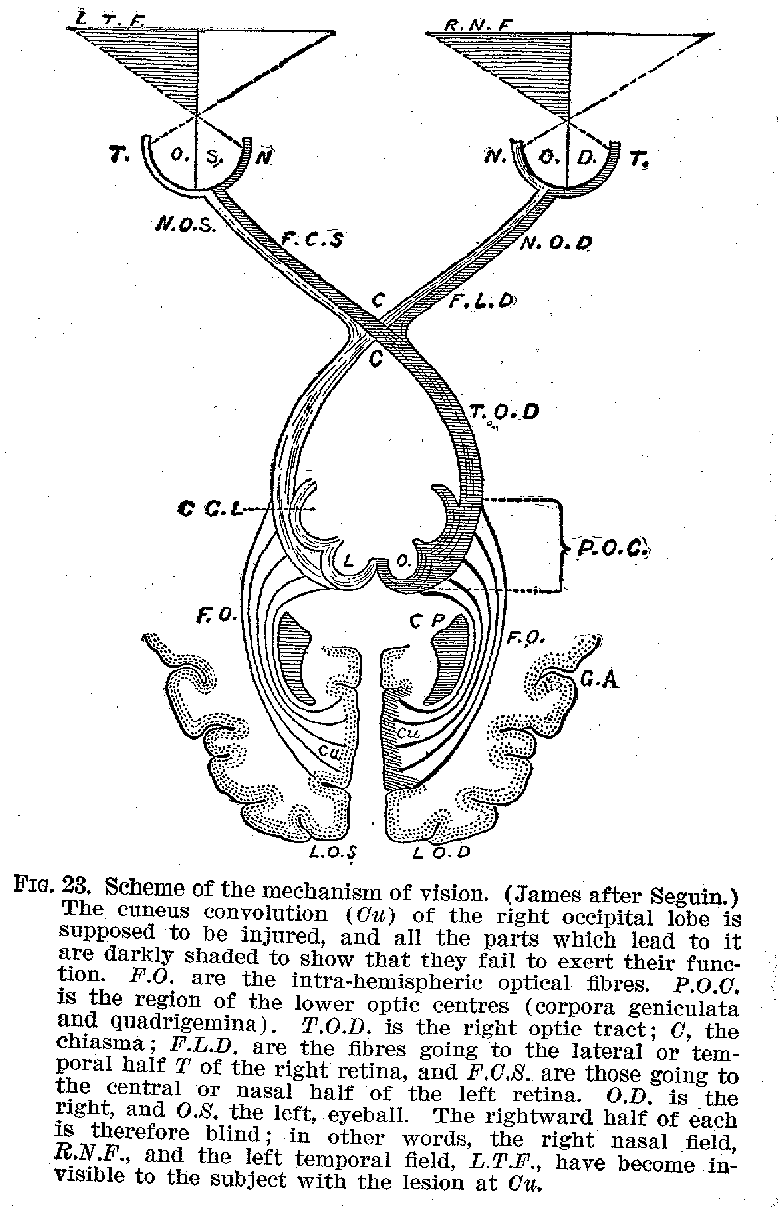
in the right side of the hemispheres. The optic nerve, however, affords a curious
modification of this plan. The neurones from the right side of each retina are connected
with the right side of the brain, those from the left side, with the left hemisphere. (See
figure 23.) In this particular, as in some others, the optic tract is peculiar. The retina
itself differs from all the other sense organs in being a part of the brain, which has in
the course of development been dislocated from its original position.
(33) and receives visual impressions. It is reasonably certain that areas
marked O receive olfactory stimuli, while the region marked B is probably that immediately
concerned with the reception of tactual and thermal stimuli. The centres for taste are not
clearly made out. It seems probable, however, that they are in the neighbourhood of the
olfactory terminals. There is reason to believe that ordinarily the peripheral sensory
neurones are in connection with the side of the cortex opposite to that from which they
originate For example, the touch nerves of the left hand find their cortical terminations
in the right side of the hemispheres. The optic nerve, however, affords a curious
modification of this plan. The neurones from the right side of each retina are connected
with the right side of the brain, those from the left side, with the left hemisphere. (See
figure 23.) In this particular, as in some others, the optic tract is peculiar. The retina
itself differs from all the other sense organs in being a part of the brain, which has in
the course of development been dislocated from its original position.
Another great group of these cortical cells in the region
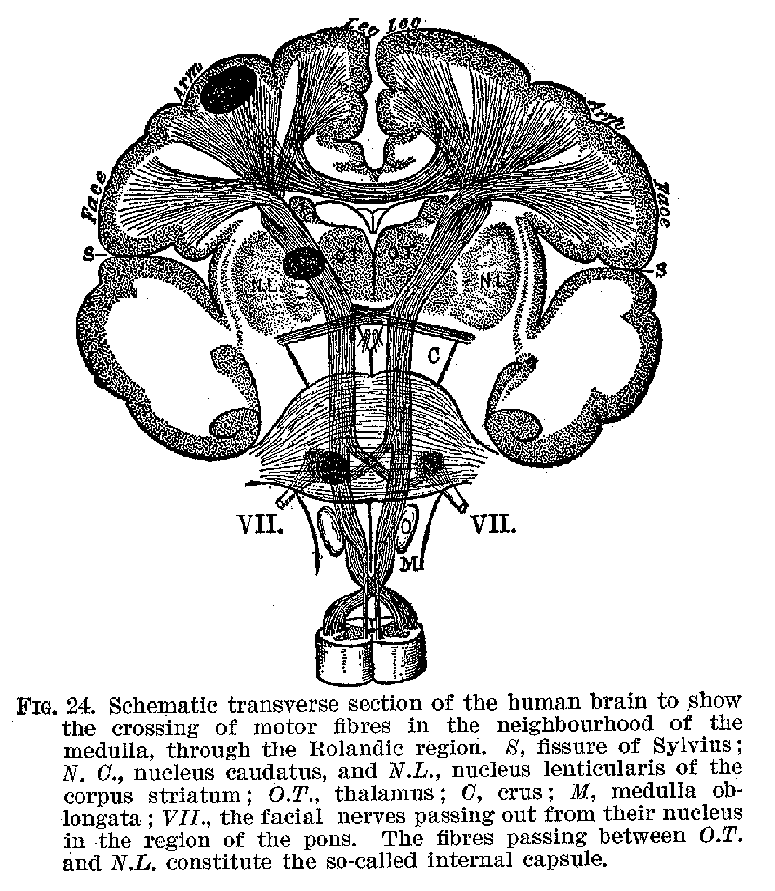
(34) marked M, generally known as the region of Rolando, from its proximity to the fissure of that name, is well recognised as being in connection with voluntary muscles, which are controlled from this centre. The voluntary muscles of each half of the body appear as a rule to be controlled by cells situated in the opposite side of the brain. (See figure 24.) In view of such facts as we have just been rehearsing, the cerebral cortex has been described as a projection system, representing every sensitive point and every voluntary muscle
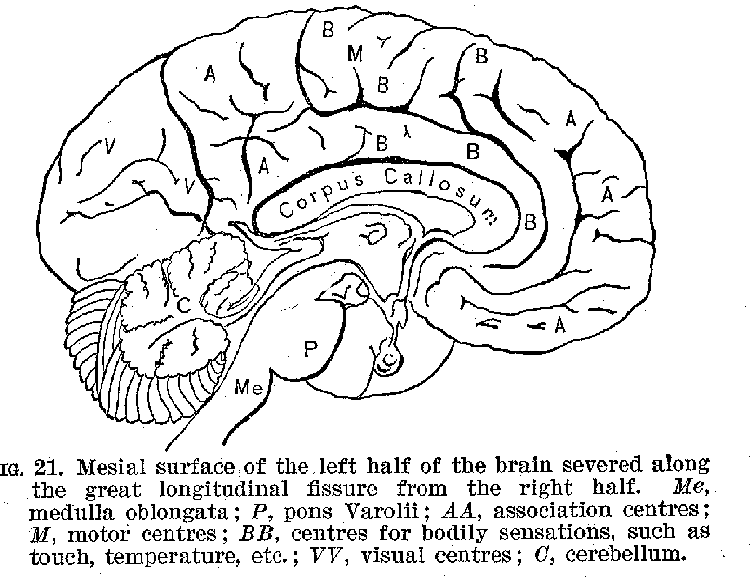
in the body. There are, however, other large areas in the cortex which are not in immediate control Of Muscles, nor do they represent the emergence point for neurones in connection with the sense organs.[2] These centres marked A are called by Flechsig, who has studied them most carefully, association centres. One of these centres, lying beneath the fissure of Sylvius, is not shown by our cuts. Their business seems to be that of uniting the several sensory regions, such as H and V, with one another and with the motor region.
(35)
(36)
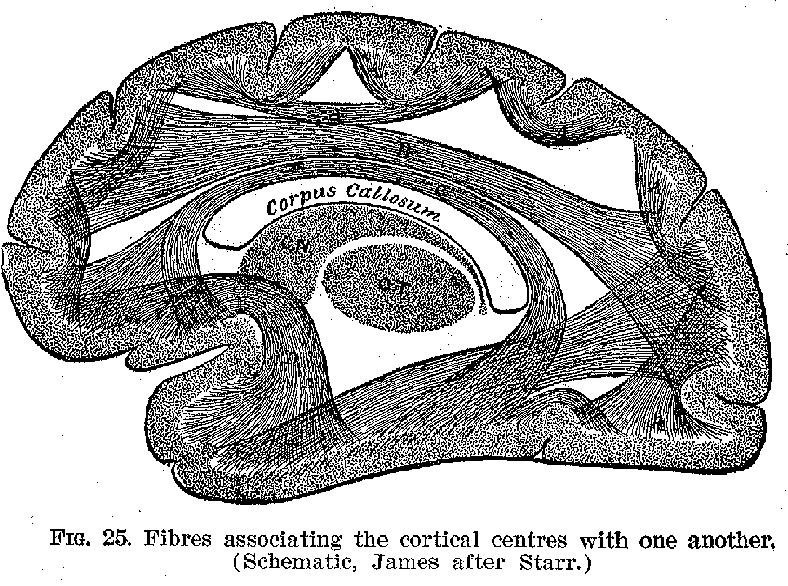
(37) (Compare figure 25.) It appears to be true in a general way at these association centres are relatively larger and more highly developed in those animals possessing most intelligence. There is another extremely important connecting mechanism, made up, however, exclusively of fibres, and
known as the corpus callosum (figures 21, 25, and 26), by means of which the two sides of the hemispheres are brought into connection with one another. These various devices make it possible for cortical nervous impulses originating in
(38) the stimulation of some sense organ, like the ear, to pass into other cortical regions like that belonging to vision, and thence out through the Rolandic zone to some muscle, producing, perhaps, a voluntary movement. This is probably
what would occur, for example, were we to bear the words "Draw a horse," then to think how a horse looks, and then finally to make the -appropriate movements of our hands. This and similar relations are suggested by figure 27.
The Cerebral Cortex and Memory -- When we contrast the cerebral cortex with the other parts of the nervous system,. with reference to its significance for consciousness, we findthat it is in the memory processes that the most conspicuous differences first come to light. If one suffers the destruction of the retina by accident or disease, or if the pathways be interrupted anywhere between the retina and the cortex, one becomes blind, but that may be all. When, however, as occasionally happens, one loses the use of the occipital regions, one may not only become blind, but one's visual memory also
 (39) is lost . It is not possible to remember how familiar objects look, and
even if seen they may not be recognized. So-called word-blindness, or visual aphasia, is
caused in this way, the patient being unable to recognise or understand written words. If
the injury is confined to one side of the brain the comm on result is hemianopsia in more
or less serious form, i. e., blindness to one-half of the field of view, owing to the
destruction of the cortical centres receiving the fibres from the lateral half of each
retina. Similarly. when the auditory region is injured, one loses the memory of auditory
experiences. If in this case, as frequently happens, the disorder be confined to one side
of the brain, and this be the side most highly developed (the left side in right-handed
people), one cannot understand what is heard. This disease is known as sensory, or
auditory, aphasia. The patient is not deaf, for the less developed and uninjured half of
the cortes may serve for the production of vague auditory consciousness, but the
associations which words and familiar sounds ordinarily evoke are wholly gone, because
these were possessions of the now diseased side. The mental condition is not unlike that
of a person hearing an unknown foreign language. He is not deaf to the words, but they
mean nothing to him, for they have no associations.
(39) is lost . It is not possible to remember how familiar objects look, and
even if seen they may not be recognized. So-called word-blindness, or visual aphasia, is
caused in this way, the patient being unable to recognise or understand written words. If
the injury is confined to one side of the brain the comm on result is hemianopsia in more
or less serious form, i. e., blindness to one-half of the field of view, owing to the
destruction of the cortical centres receiving the fibres from the lateral half of each
retina. Similarly. when the auditory region is injured, one loses the memory of auditory
experiences. If in this case, as frequently happens, the disorder be confined to one side
of the brain, and this be the side most highly developed (the left side in right-handed
people), one cannot understand what is heard. This disease is known as sensory, or
auditory, aphasia. The patient is not deaf, for the less developed and uninjured half of
the cortes may serve for the production of vague auditory consciousness, but the
associations which words and familiar sounds ordinarily evoke are wholly gone, because
these were possessions of the now diseased side. The mental condition is not unlike that
of a person hearing an unknown foreign language. He is not deaf to the words, but they
mean nothing to him, for they have no associations.
A closely comparable condition is that of motor aphasia, a disease in which one cannot articulate coherently. One is
 (40) not necessarily dumb, and there may be no true paralysis of the
articulatory muscles. But one simply cannot make the enunciatory movements in their
correct order. This disorder is often found connected in right-handed persons with disease
of the left side of the motor region of the cerebral cortex, which is in control of these
muscles. (Compare figure 19.) But it may be brought aboutand often is-as a secondary
consequence of auditory or sensory aphasia. If, when we speak, we are in the habit of
having in our minds just prior to enunciation the auditory image or thought of how the
words are going to sound, any difficulty which prevents our securing these auditory images
will effectually prevent our utterance. Now sensory aphasia involves precisely this
difficulty in commanding auditory images. As most of us do actually employ auditory
thoughts to innervate our speech muscles, for we learn to speak as children by imitating
sounds, it is surely not unnatural that sensory aphasia should so often be accompanied by
motor aphasia. Cases are on record of persons who employed visual instead of auditory
imagery to innervate the speech muscles, and who, upon suffering from lesions in the
visual regions of the brain, were seized with
(40) not necessarily dumb, and there may be no true paralysis of the
articulatory muscles. But one simply cannot make the enunciatory movements in their
correct order. This disorder is often found connected in right-handed persons with disease
of the left side of the motor region of the cerebral cortex, which is in control of these
muscles. (Compare figure 19.) But it may be brought aboutand often is-as a secondary
consequence of auditory or sensory aphasia. If, when we speak, we are in the habit of
having in our minds just prior to enunciation the auditory image or thought of how the
words are going to sound, any difficulty which prevents our securing these auditory images
will effectually prevent our utterance. Now sensory aphasia involves precisely this
difficulty in commanding auditory images. As most of us do actually employ auditory
thoughts to innervate our speech muscles, for we learn to speak as children by imitating
sounds, it is surely not unnatural that sensory aphasia should so often be accompanied by
motor aphasia. Cases are on record of persons who employed visual instead of auditory
imagery to innervate the speech muscles, and who, upon suffering from lesions in the
visual regions of the brain, were seized with
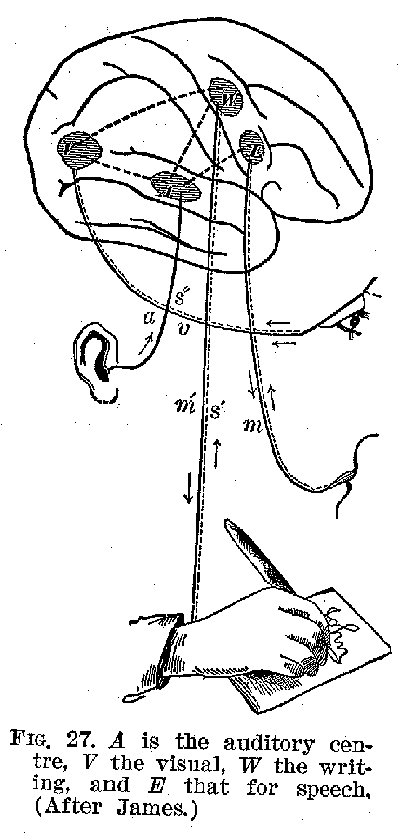
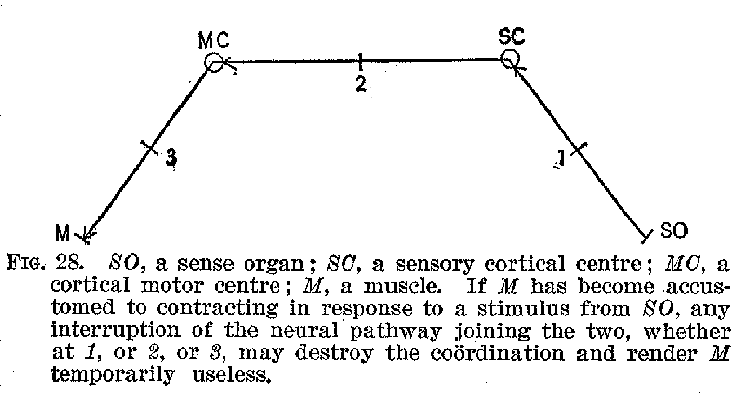
(41) motor aphasia. The sensory-motor are, or circuit, as we have previously remarked, represents the unit of action, finding ,no exception in the activity of the complex cortical centres, and any stoppage of it in the sensory portion may be as fatal to its proper operation as a defect in the muscles themselves. This is brought out in the accompanying diagram, figure 28.
Such facts as these we have been describing show us that our memory is in a peculiar way dependent upon the integrity of the cortex. Visual ideas, tactual ideas, auditory ideas, and the like can apparently be recalled only when the several parts of the cortex with which these functions are connected are intact. In the first instance a visual consciousness involves not only a visual cortex, but also a retina, and more or less of the intermediate organs between the two. A similar thing is true of the relation of all the other sense organs to the various elementary forms of sensory experience, such as touch, sound, taste, etc. But once the sensory experience has occurred, the cortex instantly takes up the impress and memory becomes possible. Destroy any part of the nervous system save this, and conscious memory may escape destruction. Destroy any specific sensory region in this cerebral cortex, and the corresponding sensory memories are obliterated or seri-
(42) -ously deranged. Destroy a region in the motor zone, and the voluntary control of some muscle, or group of muscles, is affected. Destroy or injure the association centres, and our intelligent conjoining of ideas, impressions, and movements is likely to be impaired. The gravity and permanency of these Psychical disorders brought about in the way suggested, i.e., by destruction of certain areas, varies very greatly under different conditions, so that the statements as made must be understood as attempting to convey only the broad general facts.
When one remembers that our most important and significant acts of will are based -upon hopes and fears and beliefs which involve our calling upon the memory of our past experience, one begins to appreciate how immensely important for all our life history this memory function of the cortex must be. Thus we choose, for example, one course of action rather than another, because we remember that somebody will be benefited if we act in this way, or injured if we do not. Memory always operates whenever we deliberate, and anything which would deprive us of our memory would effectually destroy the will. The cortex of the cerebrum as the physiological substrate of our conscious memory is thus the unquestioned peer among the various gross structures in the nervous system.
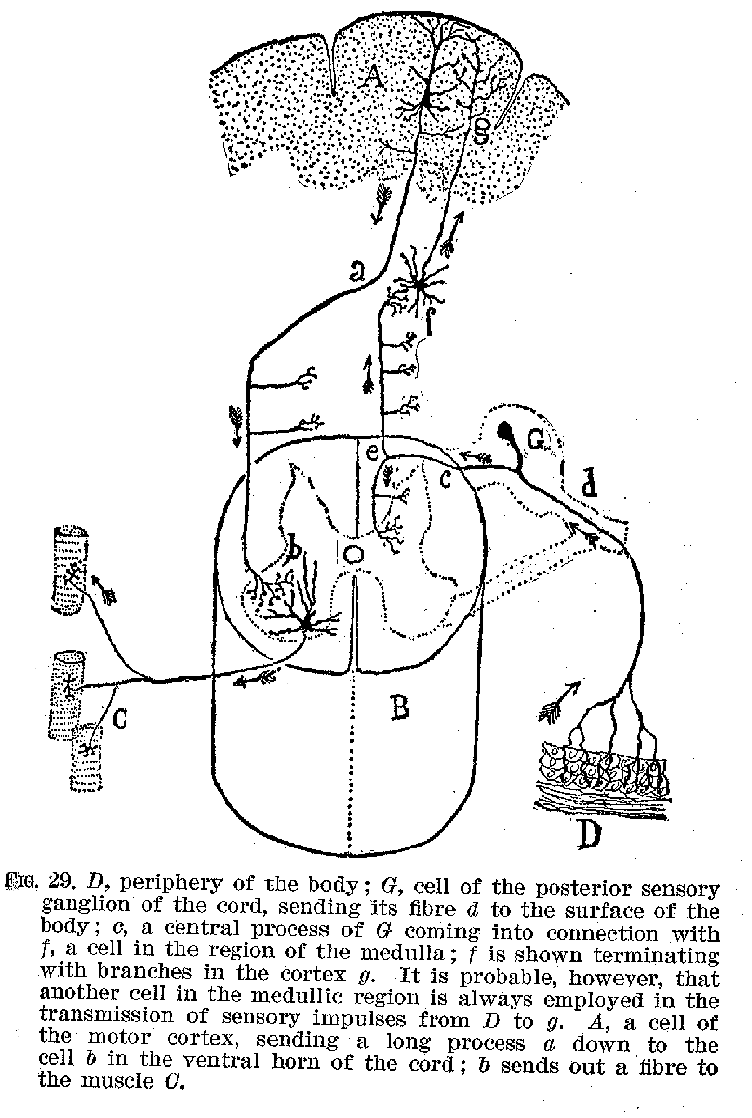
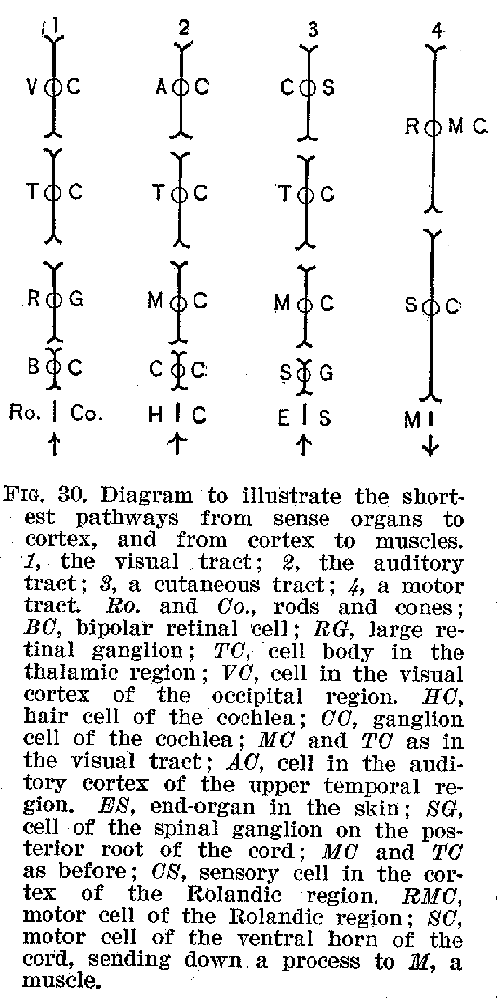
In concluding this statement, two things should be emphasised. (1) The cortex is nowhere in direct connection with a sense organ, but receives all its sensory stimulations through the intermediation of the peripheral neurones and of some of the subcortical groups, like the medulla, and it is similarly in direct connection with no voluntary muscles, but communicates with them by means of the subcortical neurones. The shortest possible pathways which could, so far as is now known, be employed in the transmission of an auditory or visual stimulus to the cortex, and back from the cortex to voluntary muscles, is shown in figures 29 and 30. The anatomical arrangements peculiar to these illustrative cases
(43) may be regarded as typical for all sense organs and voluntary muscles, but undoubtedly the pathways generally traversed nervous impulses are much more complicated and indirect.
(44)
 (2) The cortex seems always to act in an essentially unitary way.
Consciousness is, then, the counterpart of the total mass of shifting tensions going on
all over the cortex at any given mo ment. When this tension is greatest C in the occipital
re gion, we are aware of visual qualities. When the temporal convolutions are under
greatest strain, conscious ness is auditory, etc. Moreover, in this picture of con
sciousness as the counterpart of a unified series of physiological ten sions all over the
cortex, we must not forget that the whole nervous sys tem is in a measure involved. These
tensions are of such a character as to require a constant escapement through the motor
path ways, with a momentary establishment of equilibrium as a consequence of such
escapement, and a fresh disturbance of equilibrium as a secondary consequence; this latter
disturbance. being brought about
(2) The cortex seems always to act in an essentially unitary way.
Consciousness is, then, the counterpart of the total mass of shifting tensions going on
all over the cortex at any given mo ment. When this tension is greatest C in the occipital
re gion, we are aware of visual qualities. When the temporal convolutions are under
greatest strain, conscious ness is auditory, etc. Moreover, in this picture of con
sciousness as the counterpart of a unified series of physiological ten sions all over the
cortex, we must not forget that the whole nervous sys tem is in a measure involved. These
tensions are of such a character as to require a constant escapement through the motor
path ways, with a momentary establishment of equilibrium as a consequence of such
escapement, and a fresh disturbance of equilibrium as a secondary consequence; this latter
disturbance. being brought about
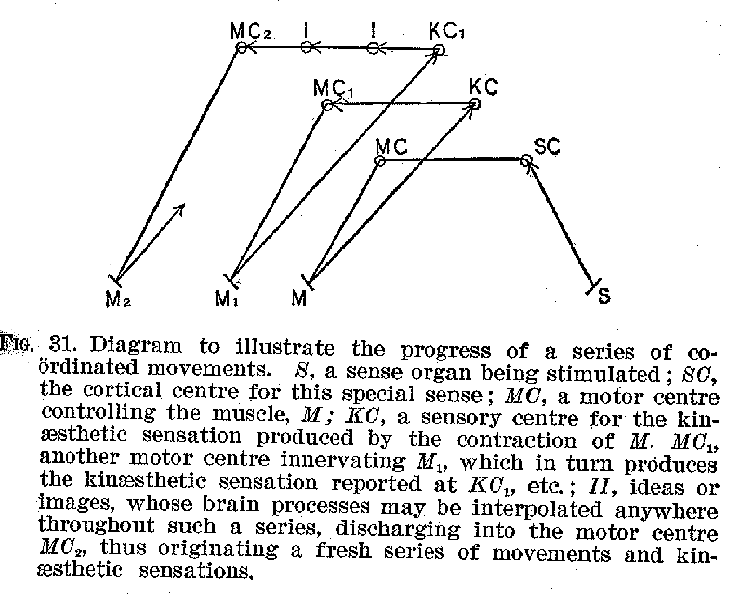
(45) through movements actually executed. Such a recurrent series of movements and sensations, illustrated by the accompanying diagram, is involved in every coherent, consecutive occupation of which we are capable. (Figure 31.) Sleep affords practically the only instance of cessation from these coordinated series of stimulations and movements. Ideational processes are often, of course, interpolated between the sensation and the movement, as is suggested by the diagram.
The Autonomic System.-- In addition to the central nervous system of which we have thus far spoken must be mentioned the autonomic system commonly known as the sympathetic system, and of which the true sympathetic is a highly important part. The autonomic system, about which our exact knowledge is lamentably defective, is apparently an outgrowth of the central system, and the two are intimately connected, both as regards their structure and their action. The striking peculiarity about the autonomic system is, as its name indicates, it, relatively self-directing or automatic activity.
(46)
The autonomic system of neurones may be conveniently, though roughly, described as made up of three great groups. One of these groups consists of a series of ganglia gathered into two long strands extending up and down each side of the spinal cord. This contains the sympathetic system in the narrower and more precise sense of the term. The second group consists of the great plexuses of ganglia found respectively in the thoracic, abdominal, and pelvic cavities. The third group consists of isolated ganglia scattered miscellaneously through the body, e. g., in the heart, in the walls of the arteries, in the eye cavity, etc. These neurone groups are made up of cell-bodies and fibres for the most part unmedullated. A portion of the impulses which affect their action apparently come from the central system. They certainly discharge impulses into the glands, the unstriped muscles, and, in the case of the heart, into striped muscle. Thus, for example, when an embarrassing announcement is made in our presence a sensory impulse passes over the auditory tract of the central system and thence, among other consequences, impulses are sent to the region in the medulla which controls the sympathetic ganglia connected with the muscular tissue of the blood-vessels, and straightway we find ourselves blushing. The sweat glands may also become active, causing us to perspire.
All the important vegetative and life sustaining processes, such as respiration, circulation, digestion, etc., are under the guidance, partial or entire, of the autonomic nerves. It is consequently to the activity of these parts that we owe our general sense of bodily well-being, as well as our feelings of distress and pain when any of these great life functions go astray. Our consciousness is undoubtedly toned, as it were, all the time, by the condition and activity of the autonomic system. This fact will become very evident when we come to study instinct and emotion. The entire nervous system, therefore, and not simply the central system, is concerned in the modifications of our consciousness.