Social Psychology
Chapter 2: The Physiological Basis of Human Behavior
Floyd Henry Allport
Table of Contents | Next | Previous
The Adaptive Function of Behavior. Behavior may be defined as the process of responding to some form of energy in the environment by an activity generally useful to life. The energy, or less exactly the object from which it is derived, is known as the `stimulus,' while the resulting activity is called the 'response.'[1] The response usually has some characteristic relation to the stimulus which evokes it, such as approaching, attacking, answering, consuming, caressing, or fleeing. The bodily structures and functions operating in behavior are of the same general sort whether the stimulus is furnished by a social or a non-social object. The first step, therefore, in the approach to social behavior is the understanding of those physiological processes involved in behavior in general.
The first stage in the adaptive process is the stimulation of the sense organ, or receptor; the final stage is the response-activity of the muscle or gland, commonly called the 'effector.' The excitation aroused in the receptor proceeds in the form of a nervous impulse along a chain of fiber-shaped nerve cells, called `neurons,' to the effector. Conduction is therefore the most elementary function of nervous tissue. The chain of neurons traversed by the impulse consists of three portions: (1) the afferent (or sensory) branch conveying the excitation from the receptor in toward the central nervous system (brain and spinal cord); (2) the central portion lying within the brain or cord and directing the impulse toward the proper outlets; and (3) the efferent (or motor) branch transmitting
( 17) the impulse outward to the effector.[2] The entire sequence is termed a reflex arc, and is to be considered as the functional unit of behavior. Especial significance attaches to the central region of the reflex arc, because it serves not only to connect the afferent and efferent portions of a single arc, but also to co�rdinate various arcs one with another. The brain and spinal cord have essentially the function of a switchboard. A complex network of millions of central neurons connects functionally each afferent pathway with every efferent and each efferent with every afferent. We receive thousands of different stimuli daily, and are capable of an enormous variety of responses. Yet so remarkable are the central adjustments between our receptors and effectors that, except in unfamiliar situations, a given stimulus almost invariably evokes the biologically correct response. The central adjustments involved in these specific responses are in some cases hereditary, and in others the result of learning through experience. In the former case they are termed reflexes, and in the latter, habits.
The Receptors and Effectors. In order to obtain a closer view of the behavior mechanism we shall discuss under separate headings the components and properties of the reflex arc and the functions of the central nervous organs. The receptors form the most natural point of departure. To do more than briefly enumerate them would, however, take us too far afield. The most important group of receptors has to do with sensations received from objects at a distance or at least external to the body, They are called the exteroceptors. The group includes the senses of vision, hearing, smell, pressure (touch), and external cold, warmth, and pain. The behavior of other persons stimulates us exclusively through the exteroceptive senses. The walls of the internal organs possess sense organs, termed interoceptors, whose stimulation gives rise to diffuse, organic, sensory experiences. Interoceptive sensations form the basis of feeling and emotion. A third group of sense organs, called proprioceptors, are embedded in the muscles, tendons, joints, anti other movable part � of the body, and are stimulated by the movements of those parts. The proprioceptive (also called
(19) �kinaesthetic') sense is necessary for learning habits and acts of skill. In this group is included the labyrinthine sense, whose receptor lies in the semicircular canals of the inner ear. Its appropriate stimulus is the rotation, movement, or change of equilibrium of the body as a whole. Proprioceptive stimulations evoke responses of movement and posture which are themselves of considerable importance as stimuli to others in social behavior.
The effectors, or organs of response, in which efferent neurons terminate, consist of muscles and glands. The two general functions of muscles are the production of movement and maintenance of posture. Muscles are attached by tendons to bones which they move on the lever principle, with joints as fulcrums, by means of contracting, that is, by shortening, the muscle fibers. The jointed parts of the body have two types of muscle, the flexor and the extensor, the former serving to bend the member at the joint, the latter to straighten it. These two types are said to be antagonistic, since in order to move the member one must be relaxed, that is, elongated, while the other is contracted. Other opposed muscle groups exist, such as the muscle lifting the eyelid and the muscle closing it, the muscles opening and closing the mouth, the circular muscles constricting (and lengthening) the intestine and the longitudinal muscle shortening it, and many others. Visceral muscles produce waves of constriction and other movements which carry on the vital processes of circulation, respiration, and digestion. Movements significant in the production of social stimuli are made chiefly by the muscles controlling the organs of speech and by those of facial expression, gesture, and bodily posture.
Glands are small, saccular, secreting organs existing either singly, as sweat and gastric glands, or grouped into complex structures, such as the thyroid, pancreas, and liver. Their secretions aid in the process of digestion and in the elimination of waste matter from the body. The so-called `ductless glands' provide internal secretions which are absorbed directly into tile blond stream. They contain substances (hormones or autacoids) which have a direct, energizing influence upon vital organs and upon bodily growth and development. Internal secretions play an important part in the emotions. The products of the sex glands are powerful
(20) internal stimulants both of sexual development and sex behavior. The secretion of tears is one of the few glandular responses of direct value as a social stimulus.
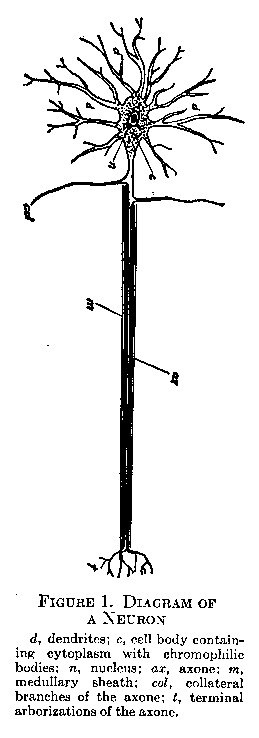
The Neuron. The nerve cells, or neurons, of which the nervous system is mainly composed, are by their structure adapted to the collection, conduction, and distribution of nervous impulses. Figure 1 shows in diagrammatic form the main features of a neuron. The cell body (c), with its nucleus (n) and flake-like masses called chromophilic bodies, is the center of the growth and nutrition of the cell. From it a group of branched fibrils, the dendrites, extend in many directions (d), and also a single fiber, usually fairly long and straight in its course, called the axone (ax). Minute thread-like structures, the neurofibrils, run freely throughout the cell body and its processes. The axones of most neurons are surrounded by a white membrane, the medullary sheath (m), which has probably an insulating and a nutritive function. Axones lying outside the brain and cord have as a further outside covering a thin sheath, the neurilemma.
Excitation in the neuron takes place at the ends of the dendrites, the impulse being received either from the specialized cells of a receptor organ, for example, the retina, or from the branched terminations of the axone of another neuron in functional connection with the first. From the dendrites the impulse is conducted through the cell body and out into the axone. The axone terminates in branched form (t in
(21) Figure I), in connection either with the dendrites of another neuron or with a muscle or gland. In afferent neurons leading from a receptor to the spinal cord the dendrite resembles an a zone in its length and medullation, the cell bode lying near the entrance of the fiber into the cord. In its function of receiving the impure it is, however, essentially a dendrite. In the brain and spinal cord there are many special types of neurons, varying in the length of their axones, and in the richness of their arborization, according to the function they fulfill.
The nature of the nerve impulse is not fully understood. It will probably be found eventually to be the propagation either of a wave of chemical reaction or of local electrical effects involving changes in the polarization of the cell membrane. The impulse does not flow as a steady stream, but as a rapid and regular succession of separate impulses forming waves of excitation along the fiber. This rhythm may be interpreted to mean that after a given impulse a certain time must elapse for the recuperation of the region of the fiber involved before it is able to transmit another impulse. The failure to respond again without an intervening period for recovery can be demonstrated by giving exceedingly rapid artificial stimulations. The interval necessary for the recovery is called the refractory period. Its length does not exceed .002 of a second. As the nerve cell recovers from the diminished excitability of the refractory period, it very shortly reaches a brief phase of hyper-excitability. If a subsequent stimuluation occurs during this phase, the amplitude of the excitation will be increased above normal. Such variations in excitability partially explain why stimuli having a certain frequency, or occurring at a certain moment, are more effective than others in producing a response, when no outward reason for such inequality of effect can be discovered. The belief is gaining ground that the strength of the nerve impulse does not vary with the intensity of the stimulus, hat 1g constant fm a given neuron. The energy of the excitation is latent in the neuron itself, and is expended in its full force or not at all. If this theory is correct, gradations in the vigor of the response must be explained by variations in the number of nerve and muscle fibers brought into play.
(22)
Reflex Arc Conduction: Properties of the Synapse. Since all reflex arcs comprise two, and most of them more than two neurons, reflex arc conduction involves the passage of the impulse from one cell to another. The finely branched terminations of an axone do not come into actual contact with the dendritic branches of the neuron next in the sequence. A minute space, filled probably by a non-neural membrane, exists between the fibrils of the two neurons. This space is called the synapse. The introduction of synapses into neural arcs has a number of important consequences which may be summarized as follows:
1. Resistance. The lengthened time required for reflex arc conduction, as compared with simple nerve fiber conduction, indicates that the synaptic gap is a region of increased resistance. 2. Polarity. The synapse serves as a kind of valve, allowing the impulse to pass only in one direction; that is, from the axone of one neuron to the dendrites of another. 3. Correlation. Synapses, being points of connection, make possible the integration of afferent neurons with a vast number of central and efferent neurons. They also permit the distribution of an impulse to a complex group of efferent pathways, such as those required in keeping one's balance on a bicycle and other acts of skill. 4. Summation. By means of the multiplicity of afferent connections afforded, synapses give opportunity for the summing up of minute and individually ineffective impulses from many neurons into an intensity of excitation sufficient to cross the threshold and be drained off into an efferent pathway. Impulses thus supplement and reinforce one another. Minute, successive, as well as simultaneous, stimuli are summed up until their total strength is sufficient to bring about a discharge across the synapse. 5. Variability of Resistance. The resistance of the synapse is altered by various conditions. Fatigue and sleep seem to increase it. It is also affected in various ways by drugs, and by changes in circulation and oxygen supply. Concurrent stimuli of a powerful character arid general nervous excitement appear to lower synaptic resistance. 6. Facilitation and Habituation. If a subject squeezes a hand-grip apparatus (dynamometer) at the time when the knee-jerk reflex is being produced by tapping the tendon below the knee, the extent of the knee-jerk will be in-
(23) -creased. This temporary effect may be interpreted either as a summation of impulses or a lowering of synaptic resistance in the knee-jerk reflex owing to an accompanying stimulus. The result, however brought about, may be regarded as the facilitation of a response by the agency of the synapse. Synaptic resistance may be decreased in a more permanent fashion by repetition. Each time a certain response is made the resistance to the impulse encountered at the synapse becomes slightly less until a complete habit is formed. 7. Inhibition. This is a necessary supplement to facilitation. Antagonistic and irrelevant responses are believed to be inhibited by an increase in their synaptic resistance, so that the response of the moment is given a `free field.' 8. Temporal Aspects. There occur at the synapse in increased form the various phases of excitability, such as refractory phase and hyper-excitability, which occur in the single nerve fiber. Professor Sherrington found that in a dog with the spinal cord severed from the brain, leaving a pure spinal reflex mechanism, the rhythmic movements of scratching could not be elicited at a more rapid rate than four per second, no matter in what rapid succession the stimuli were given. A fourth of a second, in other words, was necessary as a recuperative interval before the reflex arc was capable of functioning again. We may conclude that at their maximum rate reflexes operate in rhythms peculiar to their own refractory periods. The stage of hyper-excitability following recovery no doubt facilitates and emphasizes responses to stimuli given at the most favorable rate.
The Main Subdivisions of the Nervous System. The nervous system as a whole comprises the following subdivisions: (1) the brain; (2) the twelve pairs of cranial nerves arising from the brain, and supplying the sense organs and muscles of the head, face, and internal organs with afferent and efferent fibers; (3) the spinal cord projecting downward from the base of the brain, and protected by the bony vertebral column; (4) the spinal nerves which leave the cord in pairs at regular intervals, and furnish afferent and efferent fibers to the body wall and limbs on either side of the body; and (5) the autonomic system, an extension of the central nervous system which supplies the viscera. The first four of these parts are together called the cerebro-spinal system in distinction to the autonomic.
(24) A brief sketch of these subdivisions must suffice for our present purpose.
The Spinal Cord and Spinal Nerves. The spinal cord and brain are formed by the development and folding together of the neural groove, a depression extending lengthwise along the back of the

embryo. The closure of the sides of the groove results in a tubular structure, the hollow of which remains as the small central canal of the cord and the ventricles, or cavities, of the brain. A cross-section of the cord, as shown in Figure 2, reveals an H-shaped gray portion (G) consisting of cell bodies, dendrites, and unmedullated axone terminations; and a surrounding portion of white matter (W) made up of medullated fibers which convey impulses up or down the cord. Each spinal nerve has two roots, a dorsal root (dr), through which the afferent neurons (1) convey impulses into the posterior born of the gray matter, and a ventral root (vr) from which emerge the axones of the efferent neurons (3), their dendrites and cell bodies lying in the anterior or lateral horn of the gray matter.
The spinal cord has two important functions. The first is the conduction of afferent impulses to the brain and other levels of the central nervous system, and of efferent (motor) impulses from the brain downward to control the musculature of the limbs and trunk at various levels. The `ascending' tracts convey afferent impulses from the end organs of touch, temperature, pain, and the proprioceptive and organic senses to the higher levels of the cord and to the brain. The `descending' tracts conduct impulses downward
(25) from the brain to various spinal levels. The most important of these, the crossed and uncrossed pyramidal tracts, are composed of axones of neurons whose dendrites lie in the cerebral cortex of the brain (vide infra). These fibers all cross eventually to the side opposite to that of their cortical origin, so that the bodily movements on the left side are controlled by the right side of the brain, and those on the right side by the left side of the brain.
The second function of the cord is the correlation within itself of afferent and efferent neural pathways. A nerve impulse may come in from a receptor, for example, in the skin, and pass by one or more association (central) neurons to any efferent neuron at the same level of the cord or at a higher or lower level, and thence out to an effector. The central neuron shown in Figure 2 (2) illustrates a simple form of spinal connective. An afferent neuron, moreover, may be connected with a number of efferent neurons producing a group of serviceable co�rdinated movements without the aid of the higher centers. Spinal reflexes, however, are subject to considerable control and inhibition by impulses from the brain.
The Parts of the Brain. The brain is formed by a development of the cephalic portion of the embryonic neural tube. Beginning at the entrance of the spinal cord the main structures are the medulla, cerebellum, pons, mid-brain, and cerebral hemispheres with their basal structures the thalamus and corpus striatum. In the human brain the cerebral hemispheres are relatively very large. They overlie most of the other portions which are compactly folded beneath them and directed downward. Figure 3 represents the view of the brain which would be seen if it were divided lengthwise between the two hemispheres. It should be studied in connection with the text.
The Medulla. In the short, tapering stem by which the brain passes into the cord, known as the `medulla oblongata,' the ascending and descending tracts continue to and from the higher brain levels. The main portion of the pyramidal tracts cross in this region. Fibers of certain ascending (sensory) tracts terminate in the medulla making synaptic connections with fibers leading to the cerebellum and the cerebral hemispheres. The lower half of the series of cranial nerves rise from the medulla which thus both re-
(26) -ceives impulses from sensory surfaces of the mouth, face, and viscera, and sends motor impulses to these parts. In the medulla these regions are also brought under the control of the higher functions of the brain.
The Cerebellum, Pons, and Mid-brain. The cerebellum is a complex paired structure lying posterior to the upper part of the medulla. Its chief afferent impulses are received from the proprioceptive and labyrinthine sensory endings, and are correlated with efferent impulses which, by controlling the skeletal and trunk muscles, keep the body in equilibrium, and aid movement and co�rdination by maintaining a slight contraction (tonus) of the muscles. Tonus is observable in a bodily posture alert and prepared for action, which contrasts strongly with the weakness and inco�rdination resulting from an injury to the cerebellum. The mild afferent flow of proprioceptive impulses is released by special cerebellar, reflex mechanisms in a vigorous efferent discharge. The cerebellum is also in connection with the higher brain centers.
The pons is a transverse band of fibers passing below the medulla and connecting the right and left lobes of the cerebellum. It provides another region for conduction and for the correlation of afferent and efferent fibers in cranial nerves. The mid-brain, which serves as a stalk for the hemispheres, contains important tracts passing to and from the latter. It is also a center for reflexes controlling the eye movements, and for other visual and auditory reflexes.
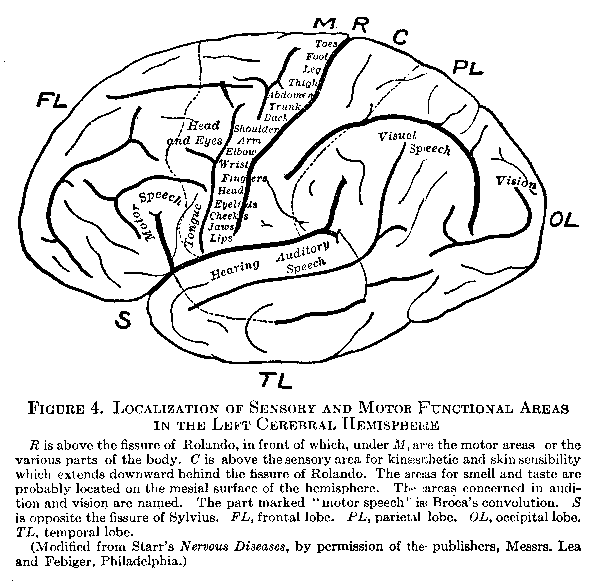
The Cerebral Hemispheres. The general appearance of the mesial and external surfaces of the cerebral hemispheres is suggested by Figures 3 and 4. Their surface consists of a layer of gray matter about four millimetres thick, called the `cortex.' The human cortex weighs only about thirteen grams, yet it contains over nine billions of nerve cells. It affords a mechanism for the correlation of impulses of almost inconceivable complexity. The cortex is greatly increased in area by being raised into folds, called `convolutions,' between which lie fissures, the two most important of which, the fissures of Rolando and Sylvius, are indicated in Figure 4 by the letters R and S. These fissures aid in dividing the cortex for purely descriptive purposes into regions called `lobes,'
(27) the most conspicuous of which (indicated in Figure 4) are the frontal, parietal, occipital, and temporal.
In the interior of the hemispheres tracts of medullated fibers, the white matter, run in various directions. They may be classified
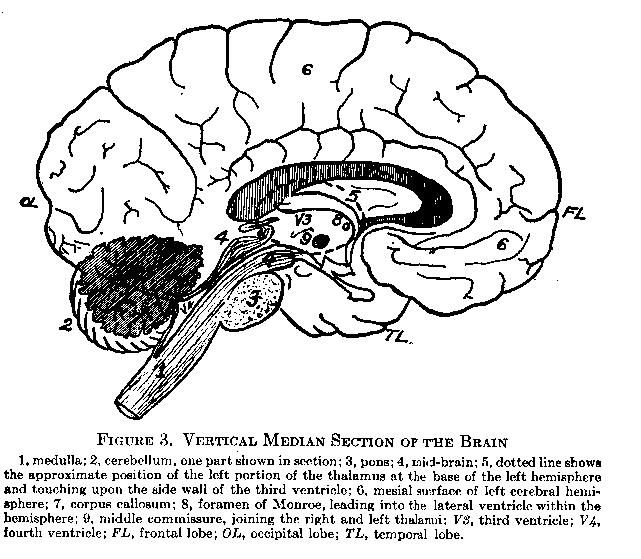
under three heads: (1) the association fibers connecting different areas of the cortex of the same hemisphere, thus serving to correlate their respective functions; (2) the commissural fibers connecting the cortices of the two hemispheres, and comprised mainly in the corpus callosum, a broad band of fibers shown in cross-section in Figure 3 (7) and (,3) the projection fibers, which extend from the cortex to lower parts of the brain, or down the spinal cord. Projection fibers are termed `ascending' or `descending' according to their function. The first pair of cranial nerves (olfactory) enters at the base of the hemispheres.
( 28)
The Thalamus and Corpus Striatum. The thalamus is a group of nerve centers of paired structure lying at the base of the cerebral hemispheres. Its position is suggested by the dotted line (5) in Figure 3. It serves as a way station for all sensory impulses (except the olfactory) arriving through the spinal and cranial nerves. Synaptic connections are here made with ascending projection fibers which spread out to all the lobes of the cortex. The thalamus serves the cortex and is subordinate to it in the following functions: (1) conduction; (2) elaboration of sensory impulses by bringing them together from various afferent channels so that they may affect the cortex in combination; (3) inhibition or blocking of irrelevant sensory stimuli (for example, pain of wounds in the emergency of battle); and (4) providing (in all probability) the neural accompaniment of states of pleasure and displeasure. The corpus striatum is a group of centers similar to the thalamus in possessing functions of scnory elaboration and sub-cortical reflexes. Its neural connection, however, with the cortex is very meager.
The Functions of the Cortex. The cortex is the chief integrating structure of the nervous system. In it areas may be distinguished having microscopical differences of cell structure which seem to signify differences of function. These areas may be grouped under the three headings — motor, sensory, and association. Figure 4 indicates their localization on the external aspect of the left hemisphere. The motor area, which lies in the oblique convolution just in front of the fissure of Rolando, contains the dendrites and cell bodies of some eighty thousand pyramidal neurons whose long axones (descending projection fibers) afford an uninterrupted conduction down the spinal cord to various levels of the trunk and limbs. The origin of the fibers controlling the various bodily regions has been localized with some exactness. The sensory areas situated in the various lobes, as indicated in Figure 4, contain the axone terminations of the neuron: constituting the final Stage in the conduction of afferent impulses from the receptors. Impulses from the auditory and optic nerves, as well as from the nerves of smell, taste, and the diffused end organs of touch, temperature, pain, and movement, are thus received in fairly distinct
(29) portions of the cortex. Each sensory area has a focalized region of pure afferent nerve endings, surrounded by a marginal area in which association fibers, communicating with other parts of the cortex, connect synaptically with the afferent terminations. The
association areas comprise extensive and complex regions of neurons lying between the areas of localized function, particularly iii the anterior frontal, parietal, and lower temporal lobes. To these areas are usually ascribed the associative processes involved in learning and thought. Developing, late both in the evolutionary : < <,11 and � in the development of the baby, they contain correlation mechanisms of a highly plastic and modifiable character.
The areas of the cortex must not be considered either as the seat of some special power or faculty, such as vision, speech, or locomo-
(30) -tion; or as the locus of the characteristic consciousness which accompanies their excitation. They are merely crucial points in the reflex arcs in which they lie. Thus the so-called `visual area' is important solely because it mediates between optical stimulations from all sorts of objects and the variety of responses by which we adjust ourselves to those objects. Our notion of mind will be clearest if we regard it neither as faculties nor states of consciousness, but as an organized system of reflex activities. Recent investigation shows that the localization of cortical functions, as suggested in Figure 4, must be accepted with strict qualifications. The cortex probably acts as a whole rather than by specific portions. Re�ducation of habits, moreover, in cases of paralysis through brain lesion proves that one part of the cortex is capable of acquiring the functions formerly possessed by another portion.
The function of cortical areas as the central portion of reflex arcs is illustrated by the defect known as aphasia. Damage to the neurons in the association areas bordering on the visual area, while it would leave the patient capable of seeing words, would render him unable to understand or to respond to them in an intelligent way. This condition is known as `word-blindness.' Word deafness is a similar defect resulting from a severance of the auditory centers from associational connections. Motor aphasia, the inability to write or speak words, though they can be perceived and understood, is due to the cutting off of the motor areas used in speaking and writing from communication with the associative functions.
Cortical Activity in Social Behavior. We may regard those parts of the central nervous system which lie below the cortex as the centers of the primitive reflex activities characteristic of all animals endowed with a nervous system. Such reflexes include respiration, digestive and excretory processes, crying, and simple movements of the appendages in defense and escape. Such reflexes are generally innate, and for their functioning the spinal cord, medulla, sere-bellum, mid-brain, and thalamus suffice. The human being, however, has added to this simple repertory a formidable array of activities whose arcs involve — and necessitate a cerebral cortex. He has acquired language, spoken and written, and other habits of
(31) skill. He has learned the use of tools, and has acquired sagacity through storing up the effects of past experiences. He has, in short, through the adaptive capacities of the cortex, attained the levels of intelligence and the power of inhibition and control which are requisite for civilized society. The chief contributions of the cortex to social behavior may be summarized as follows: (1) It underlies all solutions of human problems, which are also social problems, and makes possible their preservation in language, customs, institutions, and inventions. (2) It enables each new generation to profit by the experience of others in learning this transmitted lore of civilization. (3) It establishes habits of response in the individual for social as well as for individual ends, inhibiting and modifying primitive self-seeking reflexes into activities which adjust the individual to the social as well as to the non-social environment. Socialized behavior is thus the supreme achievement of the cortex.
The Autonomic Portion of the Nervous System. In considering the general nerve supply it is convenient to divide the body into two regions. These are the somatic, which consists of the head, trunk, limbs, and body wall, and the visceral, which includes the mouth cavity, cesophagus, stomach, intestines, lungs, heart, blood vessels, bladder, internal sex organs, and glands. The portion of the nervous system which supplies the somatic region is the cerebrospinal; the portion innervating the visceral is known as the autonomic. The autonomic is not, as its name implies, an independent system. There are no true reflexes which do not have their central portion in the cerebro-spinal axis. The autonomic system is essentially a visceral extension of the peripheral (outlying) portions of the arc. A clearer notion of the cerebro-spinal and autonomic systems may be gained if we compare their respective structure and functions. The cerebro-spinal system in the stricter sense receives its stimulations from the receptors on the surface of the body and from the. proprioceptive nerve endings. It controls skeletal and trunk muscles composed of bundles of fibers exhibiting under the miscroscope a cross-striped appearance. Its peripheral nerve fibers are medullated; and their chief function is to produce a response of movement in some part of the body, a type of muscular activity known as a 'phasic contraction.' The autonomic system has its
(32) receptors chiefly in the muscular and mucous lining of the internal organs where they are stimulated by the positions, movements, and other changes accompanying the function of these organs. The efferent autonomic fibers innervate the layers of smooth, or unstriped, muscle cells in the viscera. They control also the secretory activities of glands, including the sweat glands, as well as changes in diameter of the blood vessels and the erection of the hairs. Many autonomic fibers are unmedullated. The type of response they make in the smooth muscle is one largely of change or maintenance of muscular tension (tonus) in an organ. The pressure which an internal organ, for example, the bladder, exerts upon its contents is controlled by autonomic fibers. It is a reaction of posture rather than of movement, and is known as a tonic contraction. The central portions of the autonomic reflexes lie, as previously stated, in the brain and spinal cord.
There are three divisions of the autonomic which should be carefully distinguished. The first is the cranial division. It arises from five of the paired cranial nerves, and supplies not only certain portions of the head, such as the lens muscles and iris of the eye, and the salivary glands, but also the digestive system, bronchial tubes, and heart (see vagus nerve, Figure 5, X). In general there are interposed in the course of the efferent autonomic fibers, between their emergence from the cerebro-spinal axis and the organs they supply, certain masses of nerve centers (ganglia) lying either somewhat centrally, or (as in the case of the cranial and sacral divisions) in proximity to the organs themselves. The cranial division protects and preserves the organism by such functions as pupillary contraction and the augmenting of the glandular and muscular activities of digestion.
The second division is the sympathetic, whose distribution is illustrated in Figure 5. Its fibers emerge from the spinal nerves in the thoracic and lumbar regions (TI-TXII, LI-LV), and proceed to small segmentally arranged ganglion bodies which are vertically connected into ganglionic chains, one on either side and in front of the vertebral column. In some cases, for example, in the splanchnic nerves, the fibers run to large `collateral' ganglia of the sympathetic, not lying in the ganglionic chain itself. From these
(33)
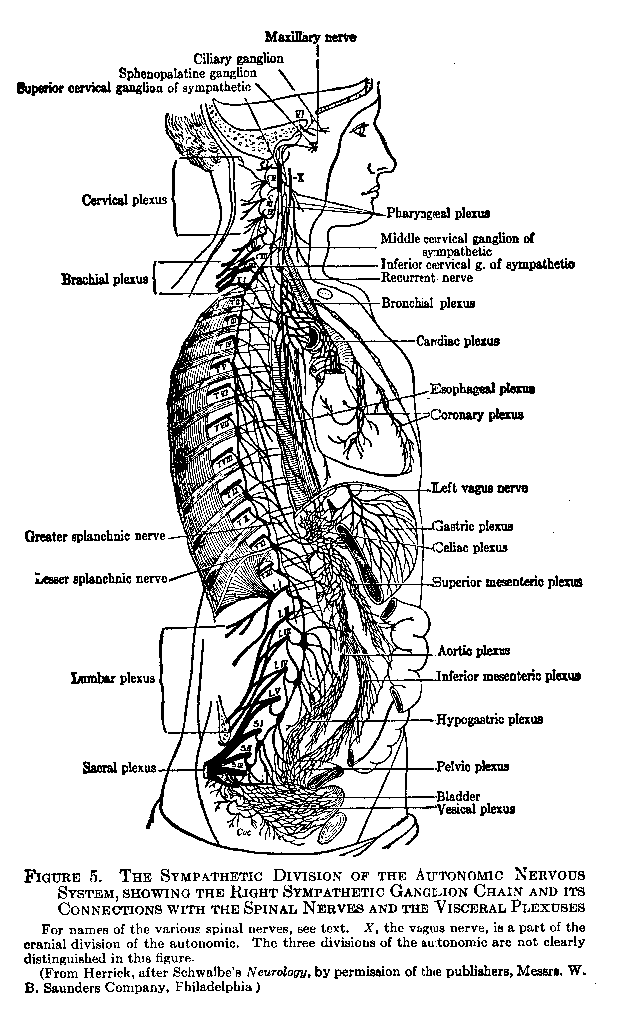
( 34) ganglia new fibers arise and run to the viscera, being distributed there in a very diffuse manner by plexuses (see Figure 5). Other fibers originating in the ganglionic chain return to the spinal nerves and are distributed to the hairs, sweat glands, and blood vessels of the outer surface of the body. The sympathetic division also supplies fibers, through ganglia in the head region, to the structures innervated by the cranial division. The sympathetic
fibers accelerate the heartbeat, constrict the blood vessels, check the contraction of the smooth muscle involved in digestion, and stop the secretion of digestive juices. Their functions are of considerable importance in emotional excitement.
( 35)
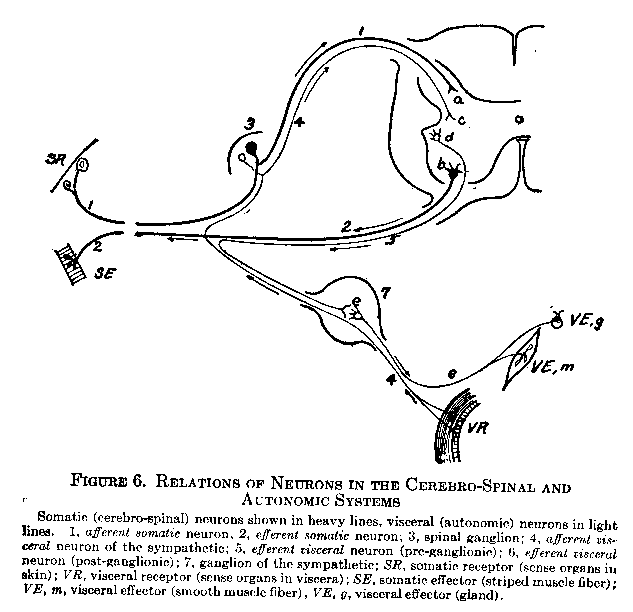
The general scheme for all autonomic innervation is as follows: An efferent autonomic, or 'visceral,' fiber leaves the cord in the ventral root of a spinal nerve, or leaves the brain in a cranial nerve, and proceeds to a ganglion. This fiber is called a pre-ganglionic neuron. Within the ganglion synaptic connection is made with a non-medullated, post-ganglionic fiber which relays the impulse to the visceral muscle or gland. The afferent visceral fiber is not broken by synapses in the ganglion body, but proceeds directly from the receptors through the dorsal root of the spinal nerve into the gray matter of the cord (through a cranial nerve to the brain in case of the cranial division). Figure 6 illustrates these connections.
The third division, the sacral, arises like the cranial directly from the cerebro-spinal axis, but at the lower end of that axis. Fibers from the sacral nerves (Figure 5, SII, iii, and iv) go directly to ganglia which supply the organs of emission — the bladder, rectum, and sexual organs. Those organs are also supplied with fibers from the sympathetic. The relations of the three divisions of the autonomic are shown diagrammatically in Figure 10 (Chapter IV).
The cranial and sacral divisions of the autonomic are often spoken of together as the cranio-sacral division. Between the functions of the sympathetic and the cranio-sacral there exists a distinct antagonism. They operate in the same organs, but with opposing reactions. For example, the sympathetic dilates the pupil, the cranial constricts it; the sympathetic inhibits digestive and sexual activities, the cranio-sacral augments them; the sympathetic accelerates the heartbeat, the cranial retards it, and so on. In addition to the three divisions of the autonomic already described there lie embedded in the walls of the heart and alimentary canal net-like plexuses of nervous tissue. These 'local plexuses' are truly autonomous. They actuate the rhythmic contractions of the viscera independently of the cerebro-spinal system which, through the three divisions of the autonomic, serves merely to regulate these functions.
The Relation of the Autonomic to the Cerebro-Spinal System. If we examine Figure 6, we shall see that the central nervous system affords a common ground for interconnection between the
( 36) cerebro-spinal reflexes of the somatic region and the autonomic reflexes of the visceral region. Thus if we imagine a central neuron connecting the various neuron endings in the cord (Figure 6), the four following possibilities arise:
1) The connection of a and b establishes a somatic-sensory, somatic-motor arc.
2) The connection of a and d establishes a somatic-sensory, visceral-motor arc.
3) The connection of c and b establishes a visceral-sensory, somatic-motor arc.
4) The connection of c and d establishes a visceral-sensory, visceral-motor arc.
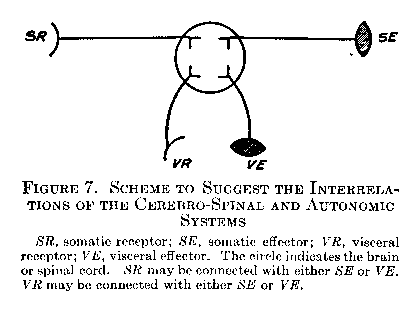
The receptor and afferent process, in other words, may be either somatic or visceral, and the efferent process and effector may be either somatic or visceral. Figure 7 illustrates the four possibilities in schematic form, and will be convenient to keep in mind as a brief summary of the behavior mechanism.
There is a tendency among those who write about the physiology of human behavior to slight the autonomic functions. From the standpoint of evolution the autonomic system is older and more fundamental than the cerebrospinal. The popular notion that the function of the stomach and other viscera is to sustain the activities of the limbs and the aesthetic and intellectual life of the `higher senses,' substitutes man's conception for Nature's. In tile light of both racial and individual development it is more correct to say that the distance receptors, the responses of locomotion, and the intellectual processes of the cortex themselves acquire their primary significance as servants of the `inner man.' Later they reach a
(37) degree of complexity which leads us to regard them as more fundamental than the visceral activities. As one writer humorously remarks, they become "the tail that wags the dog." But the autonomic functions, like still waters, run deep through life. Hunger and sexual desire, the two supreme drives of the human race, originate in autonomic receptors; while the effector side of autonomic behavior is at the base of feeling, emotion, and personality.
Compound Reflexes in Behavior. Adaptive behavior involves the extensive modification of simple innate reflexes by joining them at the synapse with other reflexes, a process which we have previously designated as correlation. A number of possibilities in the correlation of reflexes were mentioned in connection with the properties of the synapse. We have now to consider more specifically the important types of combined reflexes.
Allied and Antagonistic Reflexes. Let us suppose that one afferent neuron is in functional connection with more than one efferent, as indicated in Figure 8 (1). (Although only two efferent fibers are shown in the drawing, there may actually be many.) There are the two following possibilities: (1) The two or more effector activities may be such that they can go on simultaneously. This will be true if they do not involve antagonistic muscle groups (see p. 19). As the runner becomes `set' on the mark in response to the stimulus `ready,' all his muscles are being prepared for one concerted movement. When we greet a friend after long absence, not only the muscles employed in the handshake, but our entire effector system, takes part to some degree in the cordial response. This type of reaction is called an allied reflex. (2) The other possibility is that the effector processes may be opposed to one another. The arm cannot be both flexed and extended at the same time. A timid child pursued and tormented by an older one will sometimes turn and fight desperately, thus substituting for flight the opposed responses of attack. In this case there appears to be an inhibition at the synapse of one of the efferent pathways while the other is being employed. Reflexes of this sort are said to be mutually antagonistic.
Allied and antagonistic reflexes occur in the reverse situation; that is, where two or more afferent neurons connect with one effer-
(38) -ent, as indicated in Figure 8 (2). If the two sensory elements are. habitually associated with the same response, they will readily discharge into the common effector. An allied reflex of this sort, which is really a kind of synaptic summation, is illustrated by one's behavior in a congregation where doubt exists as to the propriety
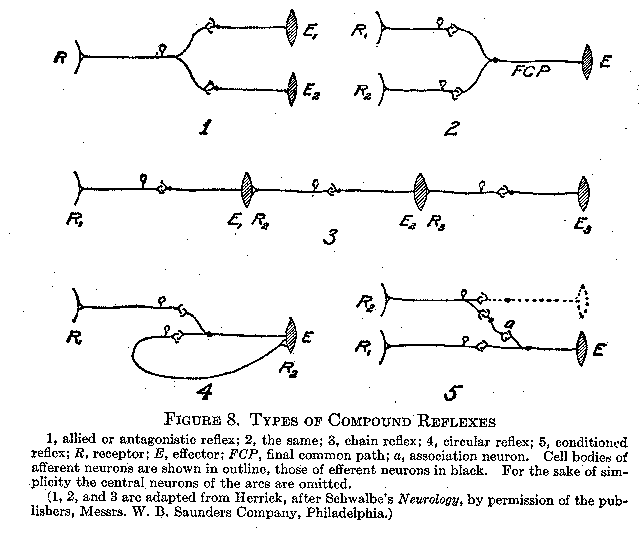
of rising on a certain occasion. The first of those to rise about one have little effect upon him, but as the numbers increase, rising becomes irresistible and automatic. When, on the other hand, the two afferent processes have paths of lowered resistance into different effectors, whose responses. arc opposed, there will result a period of indecision and delay until one reflex gains the ascendancy and the other is inhibited. The efferent outlet finally chosen is called the final common path. The behavior of two rustic characters in a well-known play illustrates this sort of antagonistic reflex in the
(39) social sphere. One had only to mention "checkers," a game in which the two were bitter rivals, and hostilities would ensue. Again, if one suggested their long comradeship and mutual sharing of joys and sorrows, they would fall into each other's arms. When the two stimuli were presented in close succession, their face depicted a period of struggle, as precarious as it was amusing, before the final common path was determined.
The Chain Reflex. Reflexes may be joined into functional patterns at the end as well as in the middle of their course. It often happens in a series of responses that one movement affords a kinaesthetic or cutaneous stimulation which evokes the next movement. In walking, the pressure on the foot and the strain on muscles, tendons, and joints resulting from one step become the stimuli for the response of taking the next step. Swallowing is a series of constrictions of the oesophagus proceeding downward by the principle of the chain reflex. Figure 8 (3) illustrates this type of reflex connection.
The Circular Reflex. The circular reflex is a special type of chain reflex in which the afferent impulse, originating from the effector response (Figure 8, 4, R2), passes back to the brain or cord and out again through the same efferent pathway previously used. The effect of this circuit is to maintain and reinforce, or to repeat, the muscular response. The holding of an object in the closed hand, and the repetition of syllables in the `talk' of the infant, probably involve this type of compound reflex. Since the response takes a rhythmic form, such synaptic conditions as length of refractory phase, hyper-excitability, and the like, are no doubt significant in its operation. The circular reflex is an indispensable aid in the infant's acquisition of speech.
The Conditioned Reflex. The most important of all the modifications of reflexes is the process by which an afferent neural pathway acquires new efferent outlets. A simple laboratory experiment will illustrate. The subject 1S seated in front of a small electrically released hammer which, by dropping close to the eye, causes him to wink. This is an original, infantile, and unconditioned reflex. A buzzer is now sounded just as, or just before, the hammer drops. The subject, of course, winks as formerly. After a number of such
(40) trials with combined stimuli, the buzzer is sounded without the release of the hammer. The subject now responds by winking to the sound of the buzzer alone. The response has been transferred or rather extended, from the biologically adequate stimulus of an object threatening the eye, to a previously ,indifferent auditory stimulus. The wink reflex may thus be said to be conditioned by the sound of the buzzer. The scheme of the conditioned reflex is shown in Figure 8 (5). R, represents the stimulation by the original stimulus, the hammer, R2 the stimulation by the `conditioning stimulus,' the buzzer, and E the eye-wink response. Through repeated joint presentation of the stimuli a path of lowered resistance is formed in some association fiber or fibers (a) already existing as a potential connection between the two reflexes.
No single law of human or animal behavior is of more far-reaching significance than that of the conditioned reflex. Half of the process of education consists of transferring appropriate responses to new and more finely discriminated stimuli. In the diagram the original response to the conditioning stimulus (for example, the buzzer) is shown in dotted lines because it is not relevant to the illustration used. In certain cases, however, the responses both to the original and conditioning stimuli are important because antagonistic; the transfer of the response from the first stimulus to the second must inhibit the original response to the latter. If, for example, a child is whipped for stealing apples, the subsequent sight of the fruit over the garden wall will of itself evoke the response of fear and avoidance aroused by the whipping with which it (the fruit) was recently associated. The original approaching response to the sight of the fruit, being antagonistic to the conditioned response, will be inhibited. We shall later observe other instances of the conditioning of original reactions through social stimuli and for the purposes of society. Recent experiments have shown that under conditions of emotional excitement, produced by autonomic responses to the stimuli employed, conditioned reflexes are formed with unusual facility. This fact is commonly recognized in the importance ascribed to such autonomically controlled factors as incentive, interest, instinctive desire, and attention. They are rightly regarded as indispensable conditions of the learning process.
(41)
The Use of the Term 'Reflex.' It should be understood that in each of the examples given above, the reaction involves not one reflex, but many. The mechanisms have been described and represented in the figures as single reflex arcs solely for the sake of clearness. Moreover, in many instances the complexities of correlation, the number of neurons involved, and the time required for the reaction, are so great that the term `reflex,' which denotes strictly only simple innate coordinations, is not applicable. This is particularly true of conditioned reactions. It would be more exact in some cases to avoid using the word `reflex,' and speak of antagonistic, allied, chain, and conditioned responses. This terminology will be employed wherever appropriate in the following chapters.
REFERENCESHerrick, C. J., An Introduction to Neurology, chs. 2-6, 12, 16, 17, 19-21. (All references are inclusive.)
Sherrington, C. S., The Integrative Action of the Nervous System, lectures 2, 4, 6, 9.
Bayliss, Wm., Principles of General Physiology (2d ed.), chs. 13, 15, 16.
Watson, J. B., Psychology from the Standpoint of a Behaviorist, chs. 2, 3, 4, 5.
Lickley, J. D., The Nervous System, chs. 2-4, 7, 9.
Dunlap, K., An Outline of Psychobiology, chs. 3, 7, 8.
Warren, H. C., Human Psychology, chs. 2-6.
Cannon, W. B., Bodily Changes in Pain, Hunger, Fear and Rage, ch. 2.
Kempf, E. J., "The Autonomic Functions and the Personality," Nervous and Mental Disease Monograph Series, no. 28, pp. 1-16.